

Substance P modulates bone remodeling properties of murine osteoblasts and osteoclasts
- PMID: 29907830
- PMCID: PMC6003941
- DOI: 10.1038/s41598-018-27432-y
Substance P modulates bone remodeling properties of murine osteoblasts and osteoclasts
Abstract
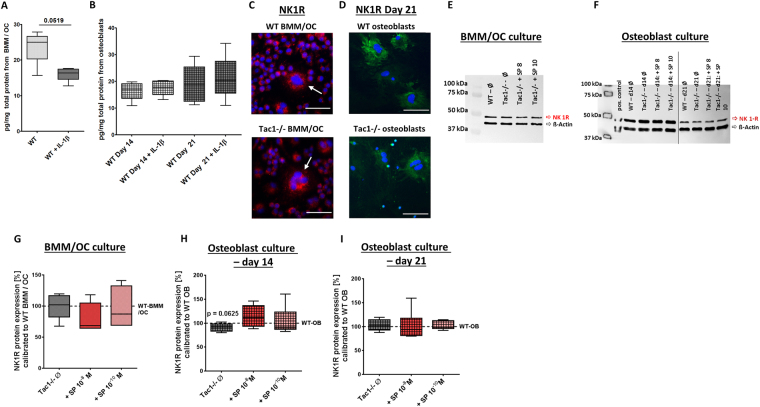
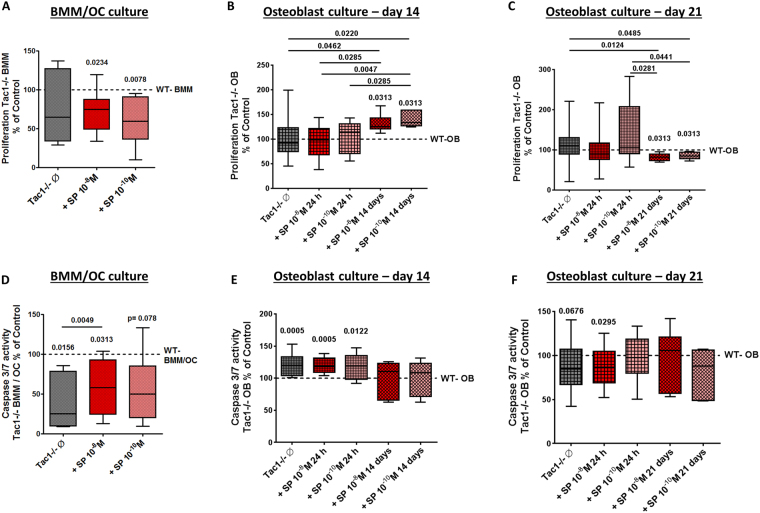
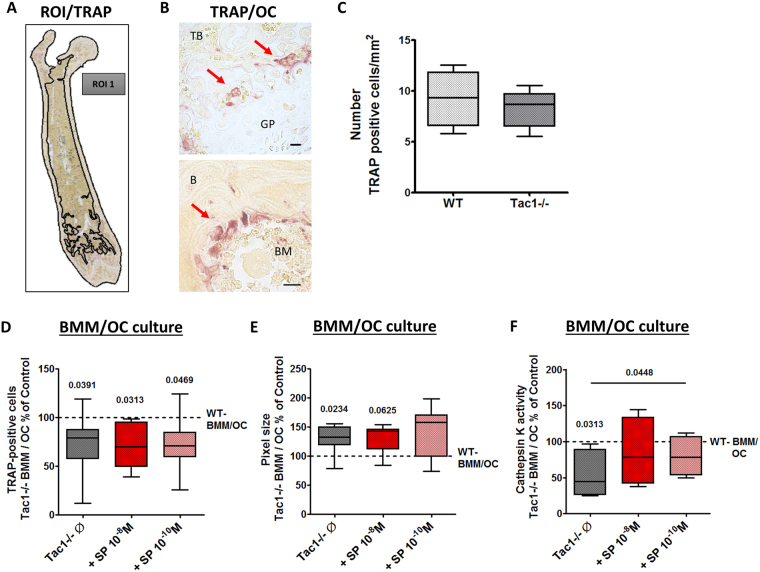
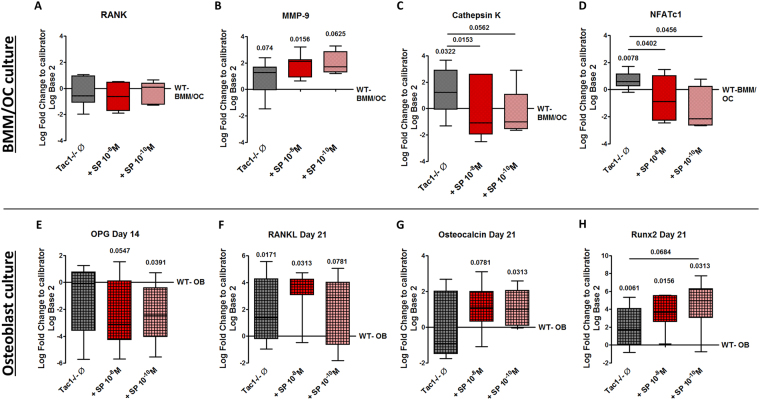
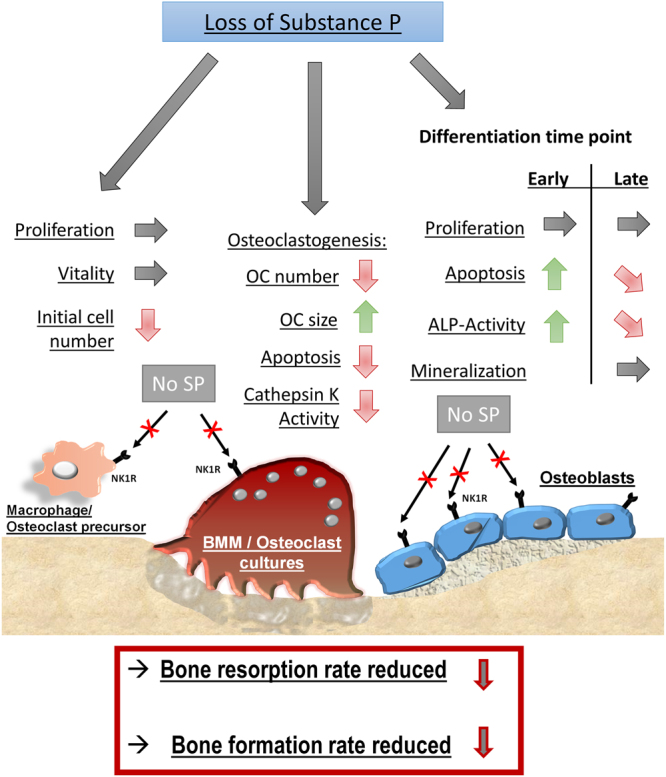
Clinical observations suggest neuronal control of bone remodeling. Sensory nerve fibers innervating bone, bone marrow and periosteum signal via neurotransmitters including substance P (SP). In previous studies we observed impaired biomechanical and structural bone parameters in tachykinin (Tac) 1-deficient mice lacking SP. Here, we aim to specify effects of SP on metabolic parameters of bone marrow macrophage (BMM)/osteoclast cultures and osteoblasts isolated from Tac1-deficient and wildtype (WT) mice. We demonstrated endogenous SP production and secretion in WT bone cells. Absence of SP reduced bone resorption rate, as we found reduced numbers of precursor cells (BMM) and multinucleated osteoclasts and measured reduced cathepsin K activity in Tac1-/- BMM/osteoclast cultures. However, this might partly be compensated by reduced apoptosis rate and increased fusion potential of Tac1-/- precursor cells to enlarged "super" osteoclasts. Contrarily, increased ALP enzyme activity and apoptosis rate during early osteoblast differentiation accelerated osteogenesis and cell death in the absence of SP together with reduced ALP activity of Tac1-/- osteoblasts during late osteogenic differentiation resulting in reduced bone formation at later stages. Therefore, we suggest that absence of SP presumably results in a slight reduction of bone resorption rate but concomitantly in a critical reduction of bone formation and mineralization rate.
Conflict of interest statement
The authors declare no competing interests.
Figures


References
-
- Raisz LG. Physiology and Pathophysiology of Bone Remodeling. Clinical Chemistry. 1999;45:1353. - PubMed
-
- Rubin, J. & Greenfield, E. In Bone Resorption Vol. 2 Topics in Bone Biology (eds Felix Bronner, MaryC Farach-Carson, & Janet Rubin) Ch. 1, 1–23 (Springer London, 2005).
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
