

Production of the compatible solute α-D-glucosylglycerol by metabolically engineered Corynebacterium glutamicum
- PMID: 29908566
- PMCID: PMC6004087
- DOI: 10.1186/s12934-018-0939-2
Production of the compatible solute α-D-glucosylglycerol by metabolically engineered Corynebacterium glutamicum
Abstract
Background: α-D-Glucosylglycerol (αGG) has beneficial functions as a moisturizing agent in cosmetics and potential as a health food material, and therapeutic agent. αGG serves as compatible solute in various halotolerant cyanobacteria such as Synechocystis sp. PCC 6803, which synthesizes αGG in a two-step reaction: The enzymatic condensation of ADP-glucose and glycerol 3-phosphate by GG-phosphate synthase (GGPS) is followed by the dephosphorylation of the intermediate by the GG-phosphate phosphatase (GGPP). The Gram-positive Corynebacterium glutamicum, an industrial workhorse for amino acid production, does not utilize αGG as a substrate and was therefore chosen for the development of a heterologous microbial production platform for αGG.
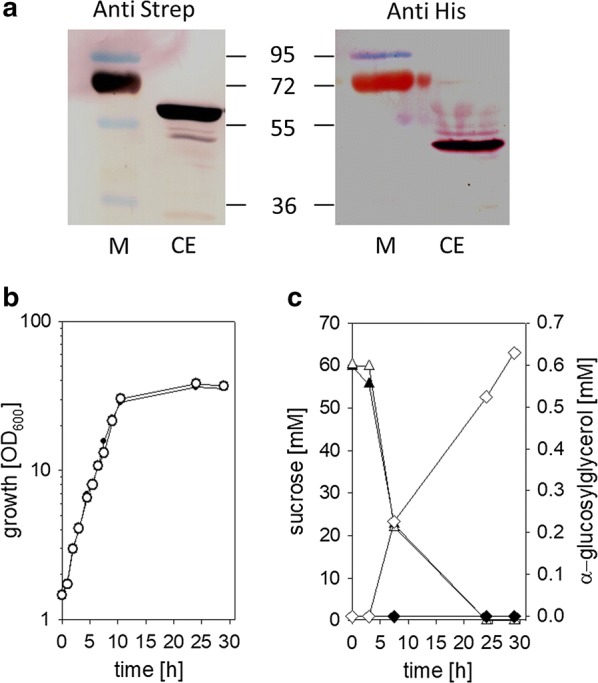
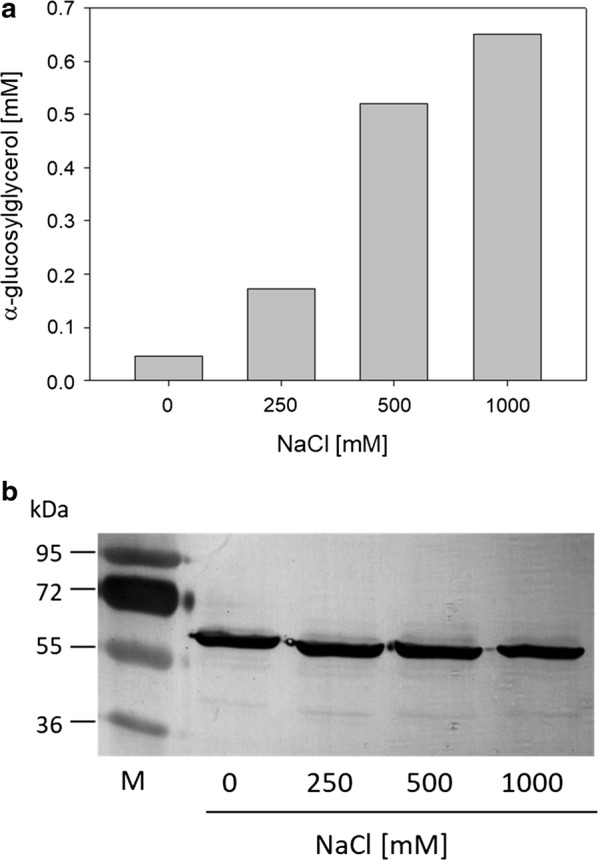
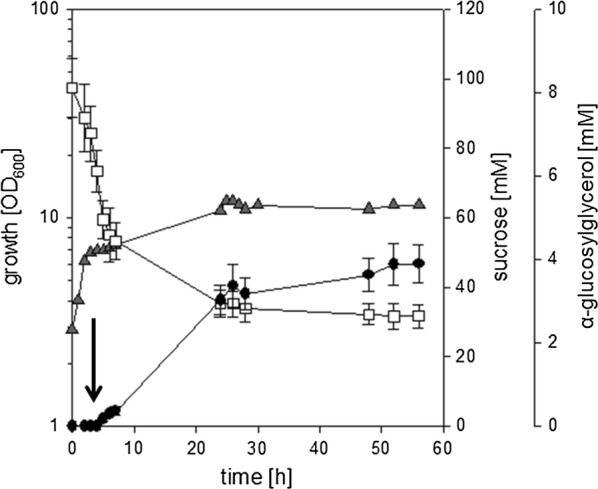
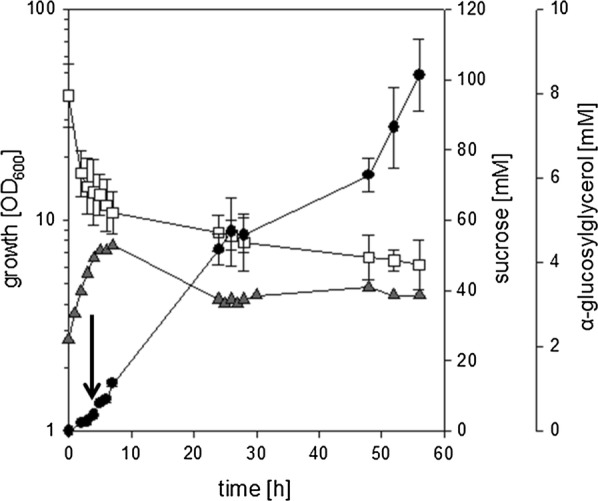
Results: Plasmid-bound expression of ggpS and ggpP from Synechocystis sp. PCC 6803 enabled αGG synthesis exclusively in osmotically stressed cells of C. glutamicum (pEKEx2-ggpSP), which is probably due to the unique intrinsic control mechanism of GGPS activity in response to intracellular ion concentrations. C. glutamicum was then engineered to optimize precursor supply for αGG production: The precursor for αGG synthesis ADP-glucose gets metabolized by both the glgA encoded glycogen synthase and the otsA encoded trehalose-6-phosphate synthase. Upon deletion of both genes the αGG concentration in culture supernatants was increased from 0.5 mM in C. glutamicum (pEKEx3-ggpSP) to 2.9 mM in C. glutamicum ΔotsA IMglgA (pEKEx3-ggpSP). Upon nitrogen limitation, which inhibits synthesis of amino acids as compatible solutes, C. glutamicum ΔotsA IMglgA (pEKEx3-ggpSP) produced more than 10 mM αGG (about 2 g L-1).
Conclusions: Corynebacterium glutamicum can be engineered as efficient platform for the production of the compatible solute αGG. Redirection of carbon flux towards αGG synthesis by elimination of the competing pathways for glycogen and trehalose synthesis as well as optimization of nitrogen supply is an efficient strategy to further optimize production of αGG.
Keywords: Compatible solute; Corynebacterium glutamicum; Glycogen; Trehalose; α-D-Glucosylglycerol.
Figures

Similar articles
-
Metabolic engineering of Corynebacterium glutamicum for enhanced production of 5-aminovaleric acid.Microb Cell Fact. 2016 Oct 7;15(1):174. doi: 10.1186/s12934-016-0566-8. Microb Cell Fact. 2016. PMID: 27717386 Free PMC article.
-
Corynebacterium glutamicum as platform for the production of hydroxybenzoic acids.Microb Cell Fact. 2018 May 12;17(1):70. doi: 10.1186/s12934-018-0923-x. Microb Cell Fact. 2018. PMID: 29753327 Free PMC article.
-
Improved fermentative production of the compatible solute ectoine by Corynebacterium glutamicum from glucose and alternative carbon sources.J Biotechnol. 2017 Sep 20;258:59-68. doi: 10.1016/j.jbiotec.2017.04.039. Epub 2017 May 3. J Biotechnol. 2017. PMID: 28478080
-
Engineering microbial cell factories: Metabolic engineering of Corynebacterium glutamicum with a focus on non-natural products.Biotechnol J. 2015 Aug;10(8):1170-84. doi: 10.1002/biot.201400590. Epub 2015 Jul 24. Biotechnol J. 2015. PMID: 26216246 Review.
-
The Actinobacterium Corynebacterium glutamicum, an Industrial Workhorse.J Microbiol Biotechnol. 2016 May 28;26(5):807-22. doi: 10.4014/jmb.1601.01053. J Microbiol Biotechnol. 2016. PMID: 26838341 Review.
Cited by
-
Salt-Regulated Accumulation of the Compatible Solutes Sucrose and Glucosylglycerol in Cyanobacteria and Its Biotechnological Potential.Front Microbiol. 2019 Sep 13;10:2139. doi: 10.3389/fmicb.2019.02139. eCollection 2019. Front Microbiol. 2019. PMID: 31572343 Free PMC article. Review.
-
The Industrial Organism Corynebacterium glutamicum Requires Mycothiol as Antioxidant to Resist Against Oxidative Stress in Bioreactor Cultivations.Antioxidants (Basel). 2020 Oct 9;9(10):969. doi: 10.3390/antiox9100969. Antioxidants (Basel). 2020. PMID: 33050339 Free PMC article.
-
Biocatalytic Production of 2-α-d-Glucosyl-glycerol for Functional Ingredient Use: Integrated Process Design and Techno-Economic Assessment.ACS Sustain Chem Eng. 2022 Jan 24;10(3):1246-1255. doi: 10.1021/acssuschemeng.1c07210. Epub 2022 Jan 11. ACS Sustain Chem Eng. 2022. PMID: 35096492 Free PMC article.
-
Continuous process technology for glucoside production from sucrose using a whole cell-derived solid catalyst of sucrose phosphorylase.Appl Microbiol Biotechnol. 2021 Jul;105(13):5383-5394. doi: 10.1007/s00253-021-11411-x. Epub 2021 Jun 30. Appl Microbiol Biotechnol. 2021. PMID: 34189615 Free PMC article.
-
Exploring the Potential of Corynebacterium glutamicum to Produce the Compatible Solute Mannosylglycerate.Front Bioeng Biotechnol. 2021 Sep 21;9:748155. doi: 10.3389/fbioe.2021.748155. eCollection 2021. Front Bioeng Biotechnol. 2021. PMID: 34621731 Free PMC article.
References
-
- da Costa MS, Santos H, Galinski EA. An overview of the role and diversity of compatible solutes in bacteria and archaea. Adv Biochem Eng Biotechnol. 1998;61:117–153. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
