

Genetic control of ColE1 plasmid stability that is independent of plasmid copy number regulation
- PMID: 29909438
- PMCID: PMC6309527
- DOI: 10.1007/s00294-018-0858-0
Genetic control of ColE1 plasmid stability that is independent of plasmid copy number regulation
Abstract
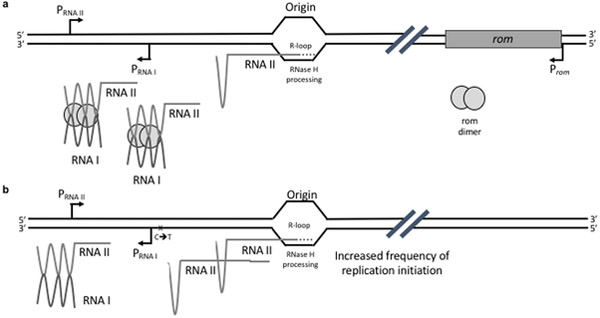
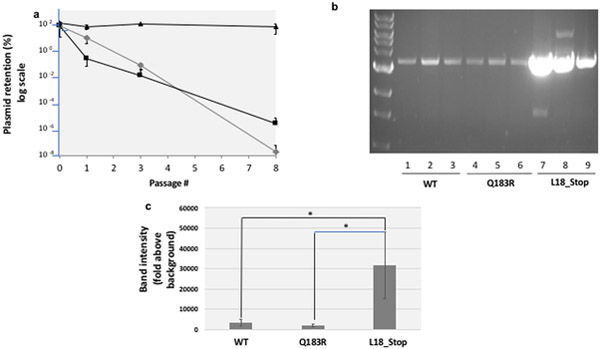
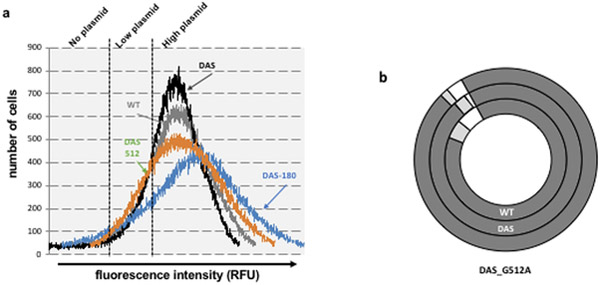
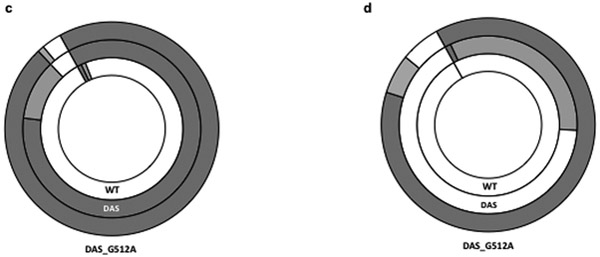
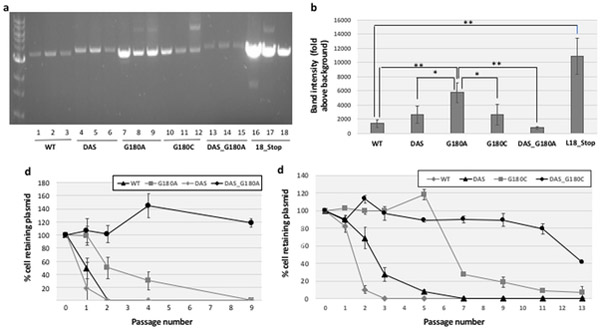
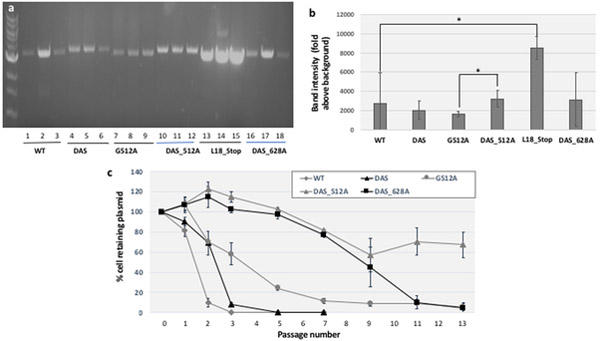
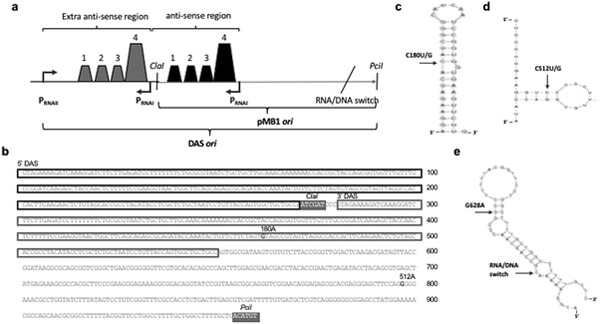
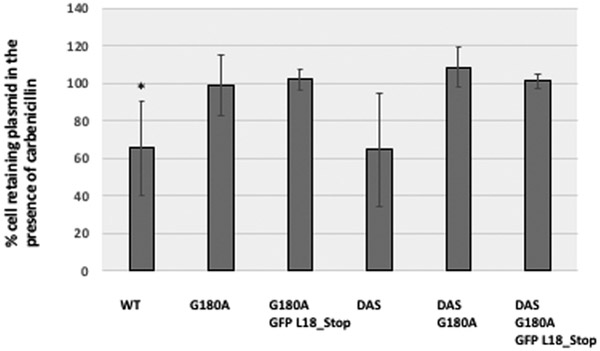
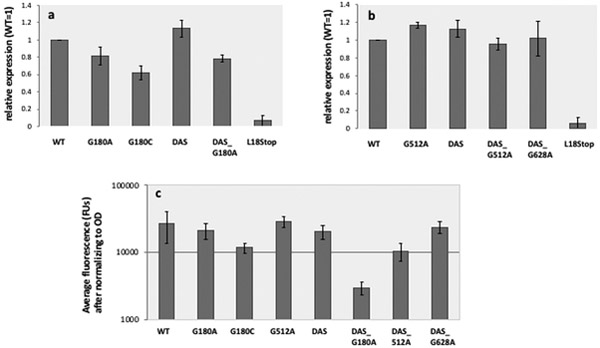
ColE1-like plasmid vectors are widely used for expression of recombinant genes in E. coli. For these vectors, segregation of individual plasmids into daughter cells during cell division appears to be random, making them susceptible to loss over time when no mechanisms ensuring their maintenance are present. Here we use the plasmid pGFPuv in a recA relA strain as a sensitized model to study factors affecting plasmid stability in the context of recombinant gene expression. We find that in this model, plasmid stability can be restored by two types of genetic modifications to the plasmid origin of replication (ori) sequence: point mutations and a novel 269 nt duplication at the 5' end of the plasmid ori, which we named DAS (duplicated anti-sense) ori. Combinations of these modifications produce a range of copy numbers and of levels of recombinant expression. In direct contradiction with the classic random distribution model, we find no correlation between increased plasmid copy number and increased plasmid stability. Increased stability cannot be explained by reduced levels of recombinant gene expression either. Our observations would be more compatible with a hybrid clustered and free-distribution model, which has been recently proposed based on detection of individual plasmids in vivo using super-resolution fluorescence microscopy. This work suggests a role for the plasmid ori in the control of segregation of ColE1 plasmids that is distinct from replication initiation, opening the door for the genetic regulation of plasmid stability as a strategy aimed at enhancing large-scale recombinant gene expression or bioremediation.
Keywords: Antisense RNA; Bioremediation; Biotechnology; ColE1 plasmid; High copy number plasmid; Origin of replication; Plasmid segregation; Plasmid stability; Recombinant expression.
Figures
References
-
- Brantl S (2014). Plasmid Replication Control by Antisense RNAs. Microbiology spectrum 2, PLAS-0001–2013. - PubMed
-
- Cesareni G, Helmer-Citterich M, and Castagnoli L (1991). Control of ColE1 plasmid replication by antisense RNA. Trends Genet 7, 230–235. - PubMed
-
- Cooper NS, Brown ME, and Caulcott CA (1987). A mathematical method for analysing plasmid stability in micro-organisms. J Gen Microbiol 133, 1871–1880. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
