

A Single-Cell Transcriptome Atlas of the Aging Drosophila Brain
- PMID: 29909982
- PMCID: PMC6086935
- DOI: 10.1016/j.cell.2018.05.057
A Single-Cell Transcriptome Atlas of the Aging Drosophila Brain
Abstract
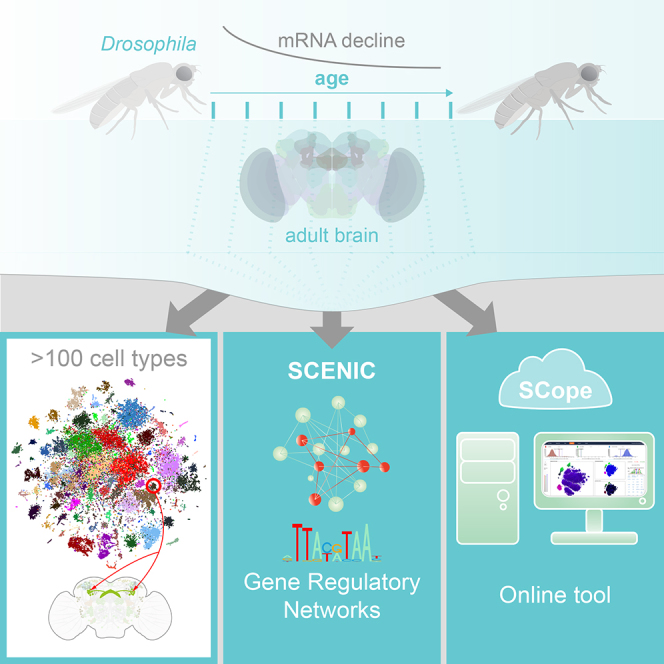
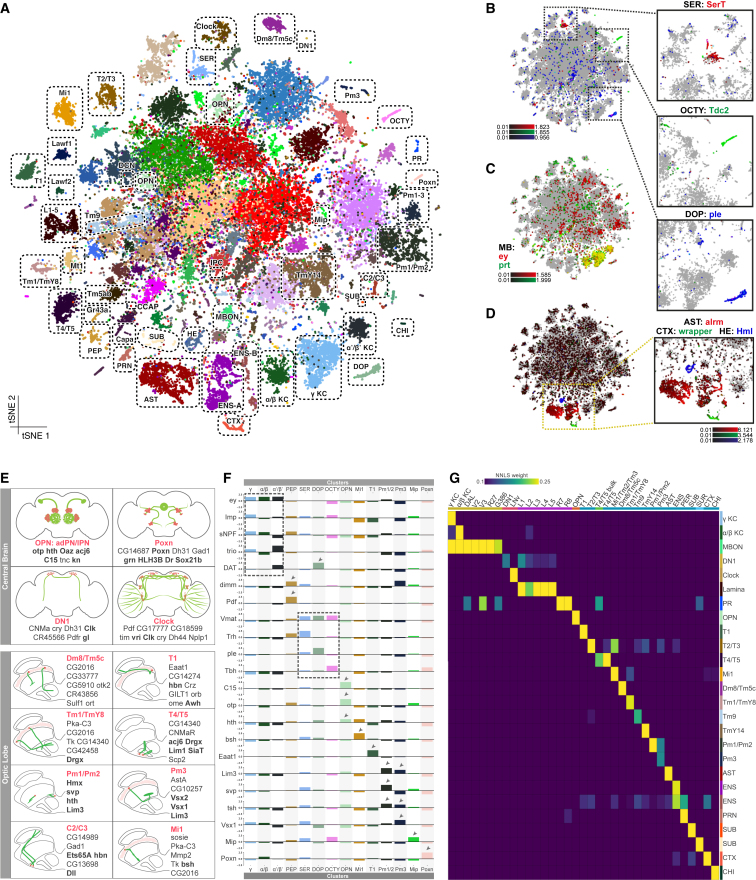
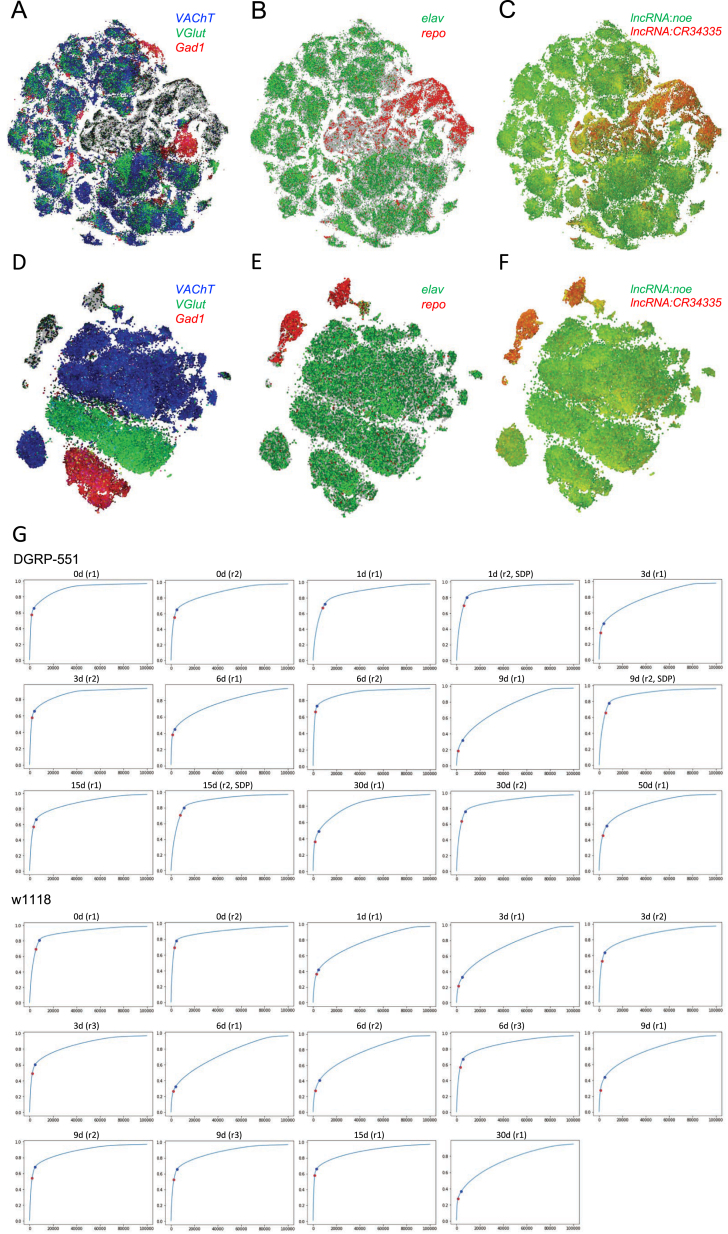
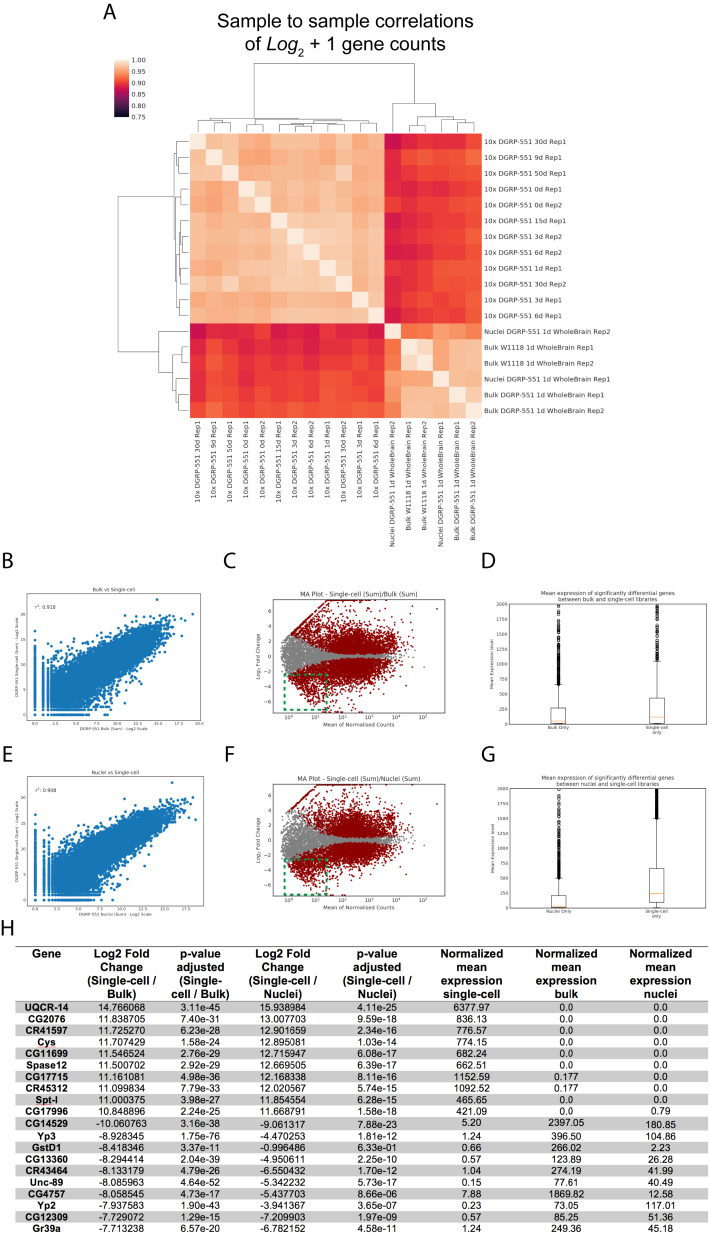
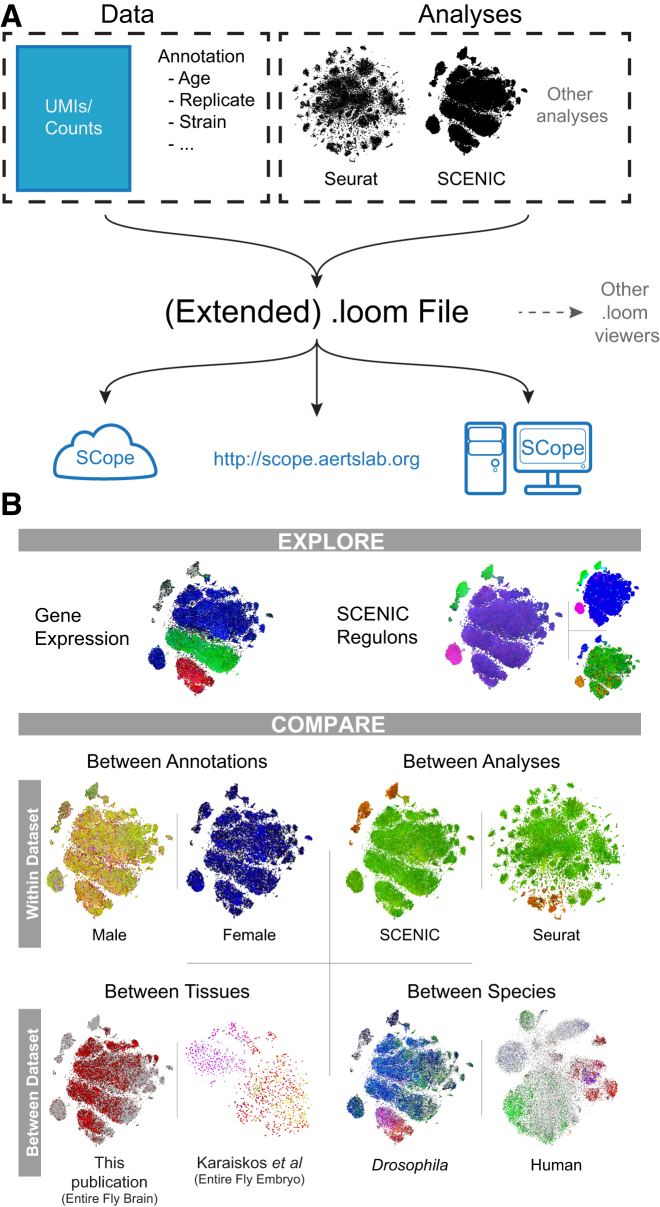
The diversity of cell types and regulatory states in the brain, and how these change during aging, remains largely unknown. We present a single-cell transcriptome atlas of the entire adult Drosophila melanogaster brain sampled across its lifespan. Cell clustering identified 87 initial cell clusters that are further subclustered and validated by targeted cell-sorting. Our data show high granularity and identify a wide range of cell types. Gene network analyses using SCENIC revealed regulatory heterogeneity linked to energy consumption. During aging, RNA content declines exponentially without affecting neuronal identity in old brains. This single-cell brain atlas covers nearly all cells in the normal brain and provides the tools to study cellular diversity alongside other Drosophila and mammalian single-cell datasets in our unique single-cell analysis platform: SCope (http://scope.aertslab.org). These results, together with SCope, allow comprehensive exploration of all transcriptional states of an entire aging brain.
Keywords: Drosophila; aging; brain; gene regulatory networks; mitochondria; neuronal subtypes; oxidative phosphorylation; single-cell RNA-seq; single-cell bioinformatics.
Copyright © 2018 The Authors. Published by Elsevier Inc. All rights reserved.
Figures

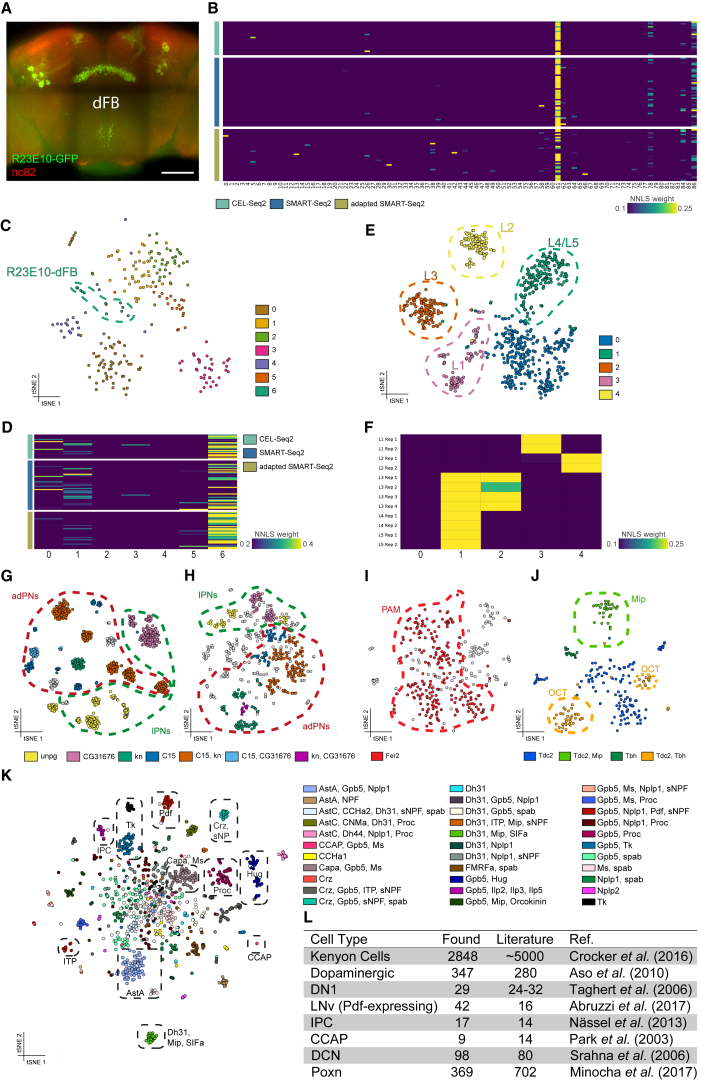
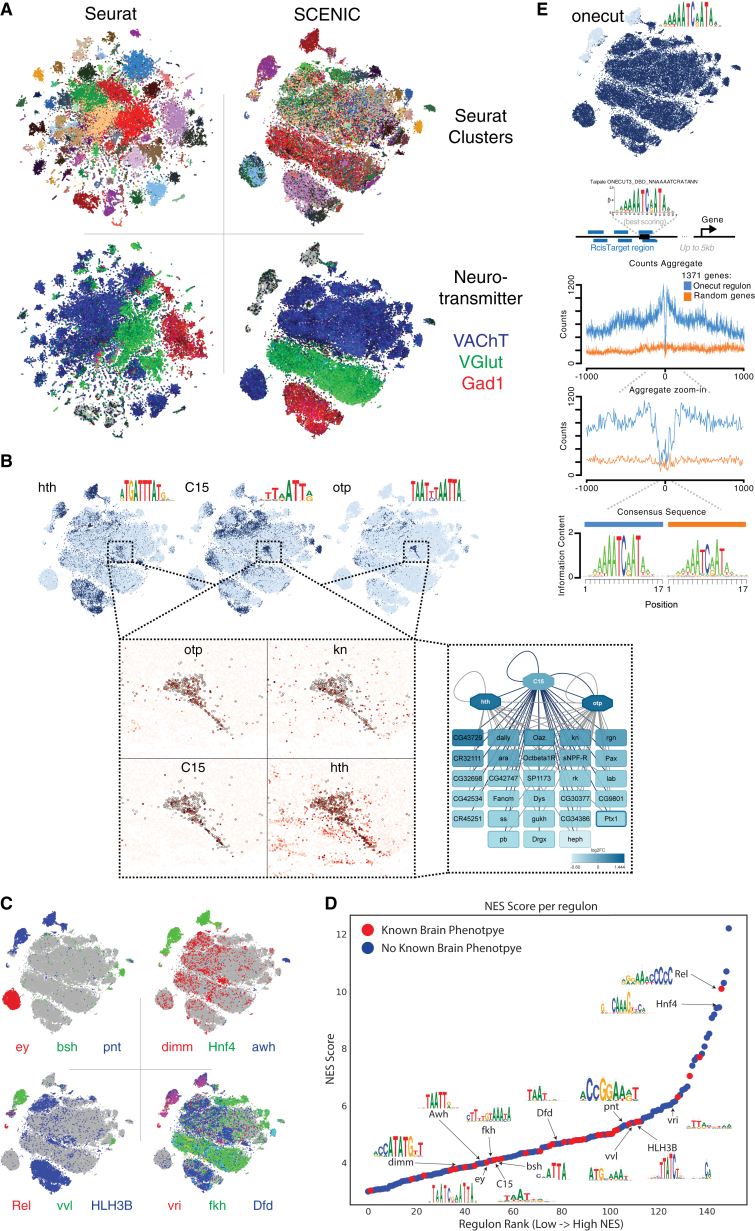
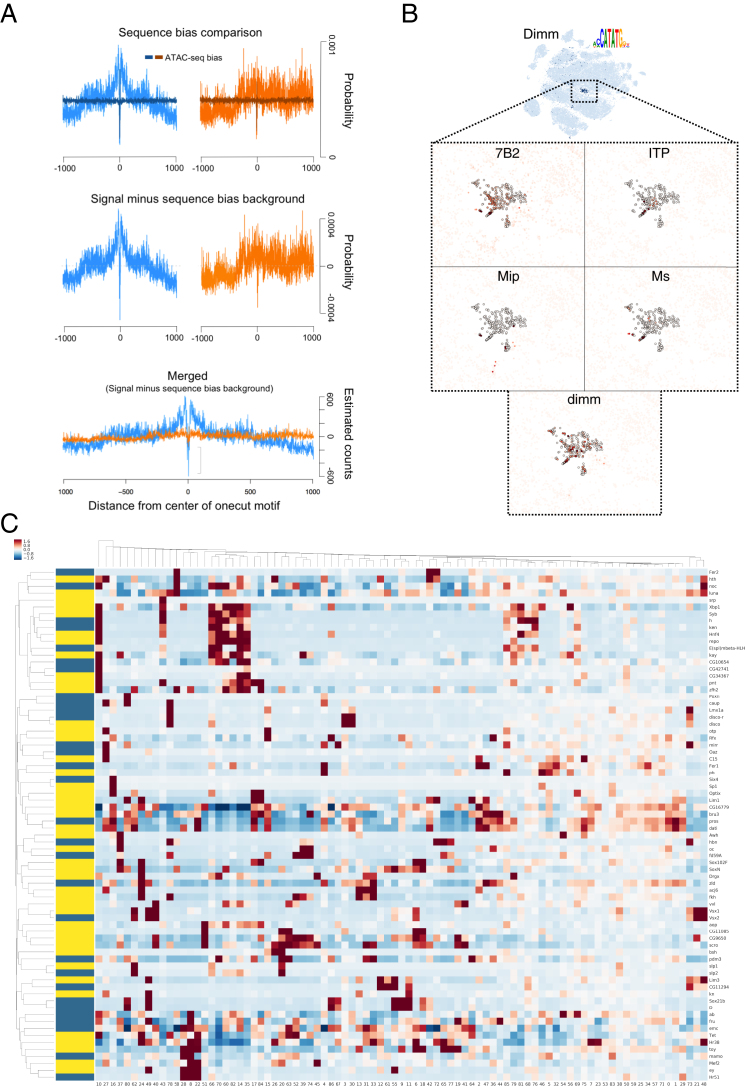
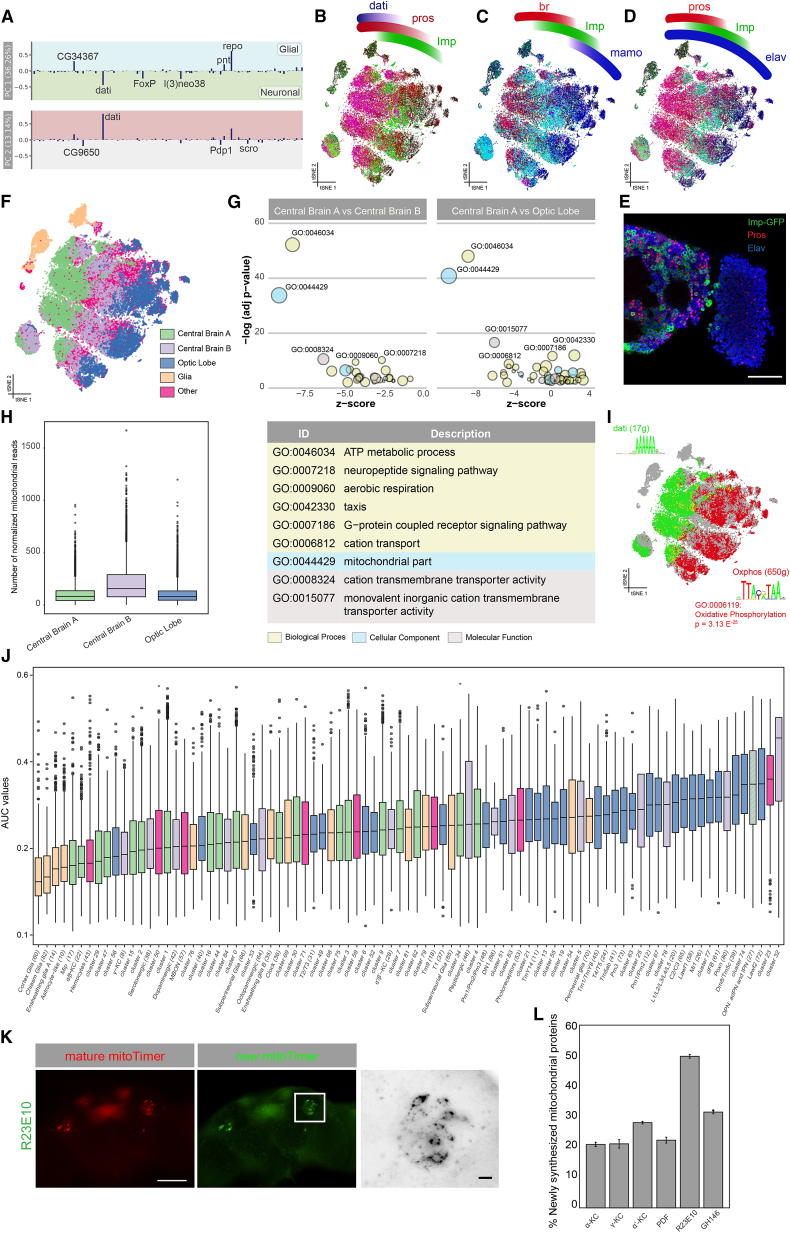
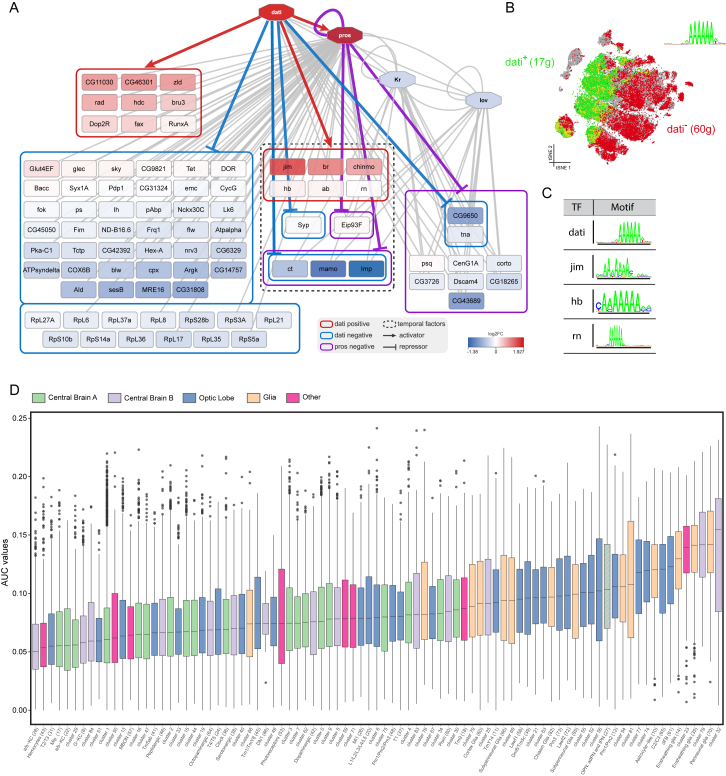
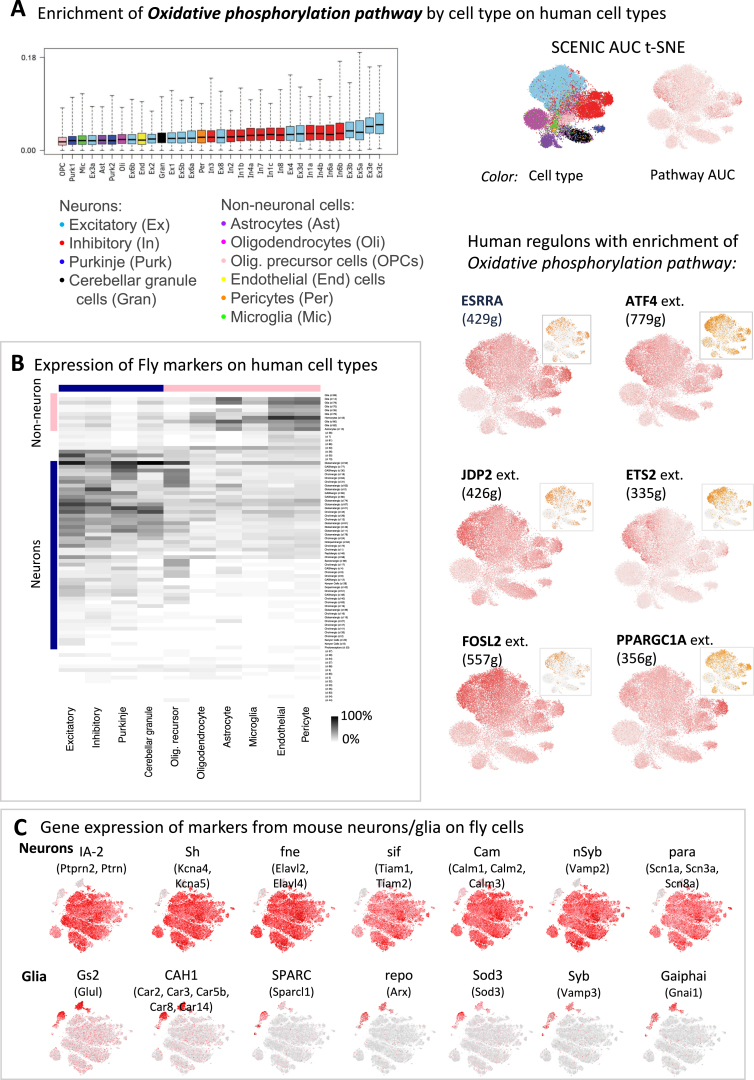

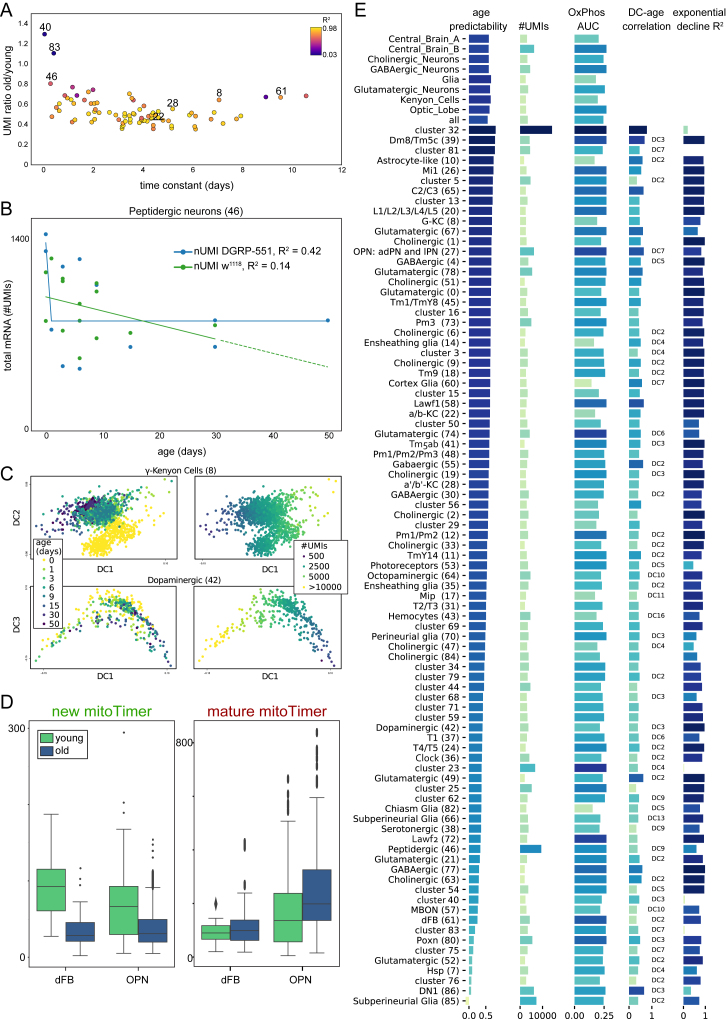
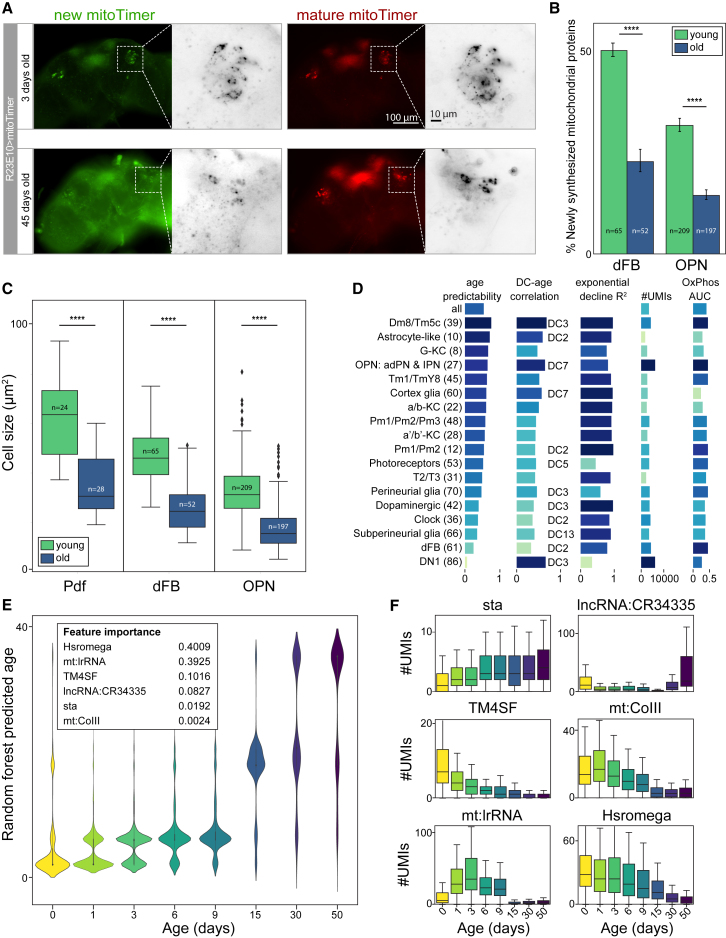
Comment in
-
Sequencing Diversity One Cell at a Time.Cell. 2018 Aug 9;174(4):777-779. doi: 10.1016/j.cell.2018.07.024. Cell. 2018. PMID: 30096308
References
-
- Angerer P., Haghverdi L., Büttner M., Theis F.J., Marr C., Buettner F. destiny: diffusion maps for large-scale single-cell data in R. Bioinformatics. 2016;32:1241–1243. - PubMed
-
- Aronesty E. Expression Analysis; Durham, NC: 2011. ea-utils: Command-line tools for processing biological sequencing data.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
