

Reconfiguration of a Multi-oscillator Network by Light in the Drosophila Circadian Clock
- PMID: 29910074
- PMCID: PMC6039274
- DOI: 10.1016/j.cub.2018.04.064
Reconfiguration of a Multi-oscillator Network by Light in the Drosophila Circadian Clock
Abstract
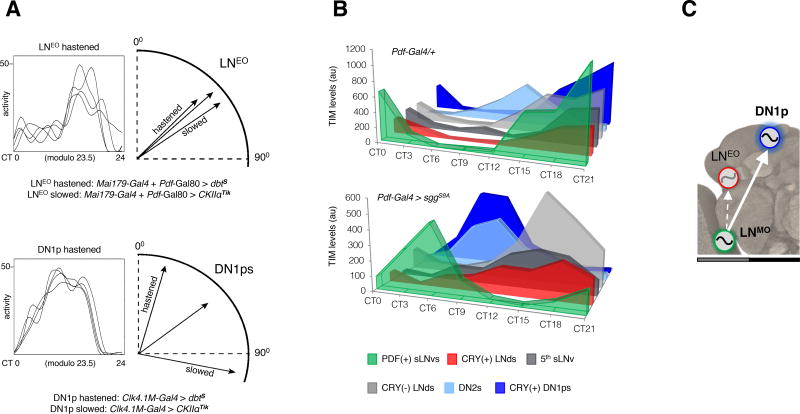
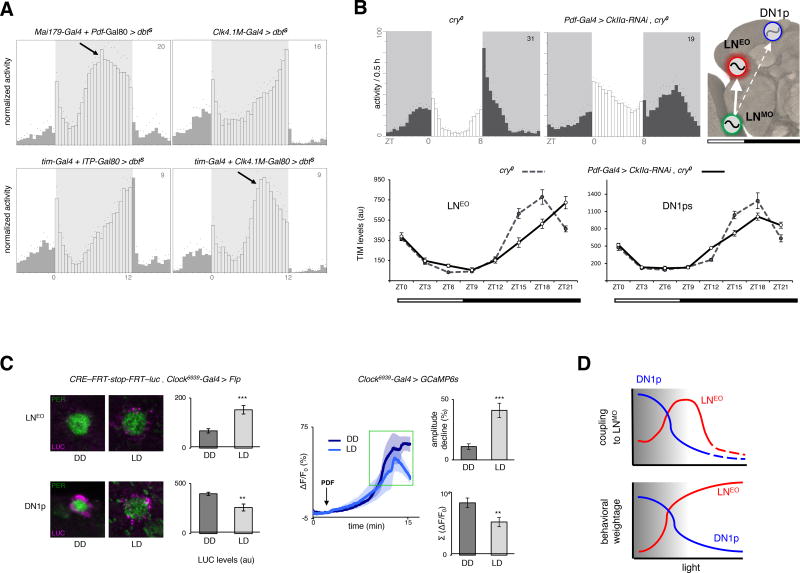
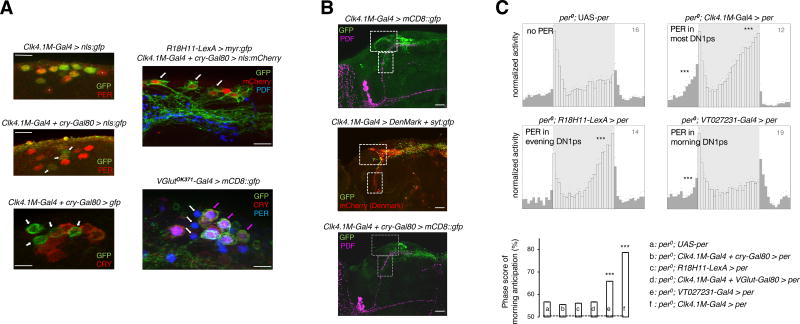
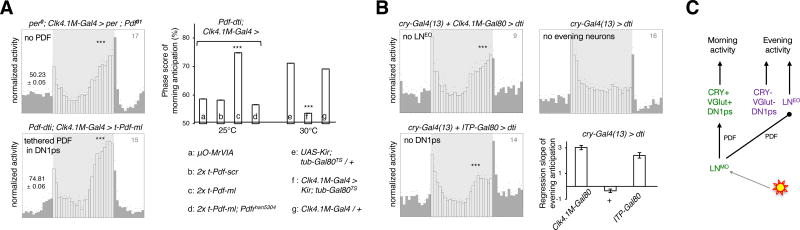
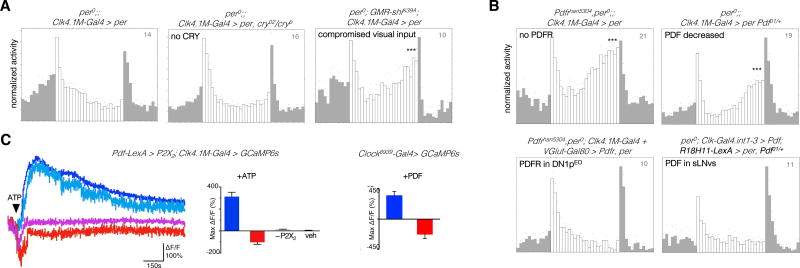
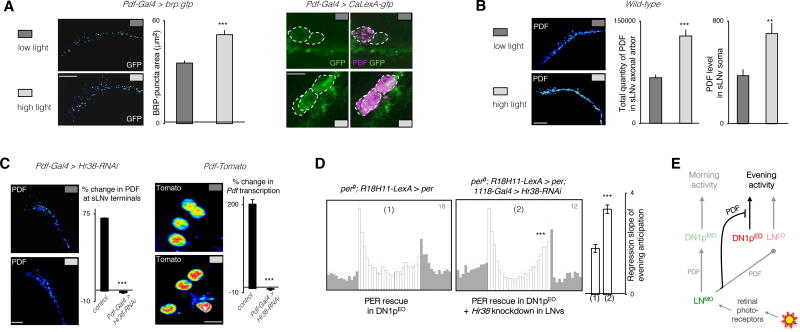
The brain clock that drives circadian rhythms of locomotor activity relies on a multi-oscillator neuronal network. In addition to synchronizing the clock with day-night cycles, light also reformats the clock-driven daily activity pattern. How changes in lighting conditions modify the contribution of the different oscillators to remodel the daily activity pattern remains largely unknown. Our data in Drosophila indicate that light readjusts the interactions between oscillators through two different modes. We show that a morning s-LNv > DN1p circuit works in series, whereas two parallel evening circuits are contributed by LNds and other DN1ps. Based on the photic context, the master pacemaker in the s-LNv neurons swaps its enslaved partner-oscillator-LNd in the presence of light or DN1p in the absence of light-to always link up with the most influential phase-determining oscillator. When exposure to light further increases, the light-activated LNd pacemaker becomes independent by decoupling from the s-LNvs. The calibration of coupling by light is layered on a clock-independent network interaction wherein light upregulates the expression of the PDF neuropeptide in the s-LNvs, which inhibits the behavioral output of the DN1p evening oscillator. Thus, light modifies inter-oscillator coupling and clock-independent output-gating to achieve flexibility in the network. It is likely that the light-induced changes in the Drosophila brain circadian network could reveal general principles of adapting to varying environmental cues in any neuronal multi-oscillator system.
Keywords: Drosophila; circadian clock; hormone receptor-like in 38; light; morning and evening oscillators; oscillator coupling; pigment-dispersing factor; posterior dorsal neurons 1; rest-activity rhythms; visual system.
Copyright © 2018 Elsevier Ltd. All rights reserved.
Figures
References
-
- Beckwith EJ, Ceriani MF. Communication between circadian clusters: The key to a plastic network. FEBS Lett. 2015;589:3336–3342. - PubMed
-
- Yoshii T, Rieger D, Helfrich-Forster C. Two clocks in the brain: An update of the morning and evening oscillator model in Drosophila. Prog Brain Res. 2012;199:59–82. - PubMed
-
- Lucas RJ. Mammalian inner retinal photoreception. Curr Biol. 2013;23:R125–33. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
