

Archaeal cell surface biogenesis
- PMID: 29912330
- PMCID: PMC6098224
- DOI: 10.1093/femsre/fuy027
Archaeal cell surface biogenesis
Abstract
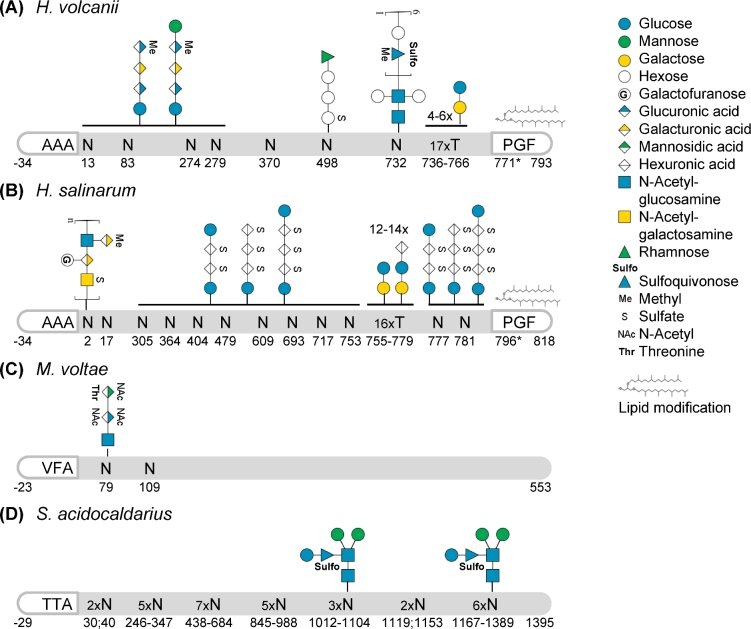
Cell surfaces are critical for diverse functions across all domains of life, from cell-cell communication and nutrient uptake to cell stability and surface attachment. While certain aspects of the mechanisms supporting the biosynthesis of the archaeal cell surface are unique, likely due to important differences in cell surface compositions between domains, others are shared with bacteria or eukaryotes or both. Based on recent studies completed on a phylogenetically diverse array of archaea, from a wide variety of habitats, here we discuss advances in the characterization of mechanisms underpinning archaeal cell surface biogenesis. These include those facilitating co- and post-translational protein targeting to the cell surface, transport into and across the archaeal lipid membrane, and protein anchoring strategies. We also discuss, in some detail, the assembly of specific cell surface structures, such as the archaeal S-layer and the type IV pili. We will highlight the importance of post-translational protein modifications, such as lipid attachment and glycosylation, in the biosynthesis as well as the regulation of the functions of these cell surface structures and present the differences and similarities in the biogenesis of type IV pili across prokaryotic domains.
Figures
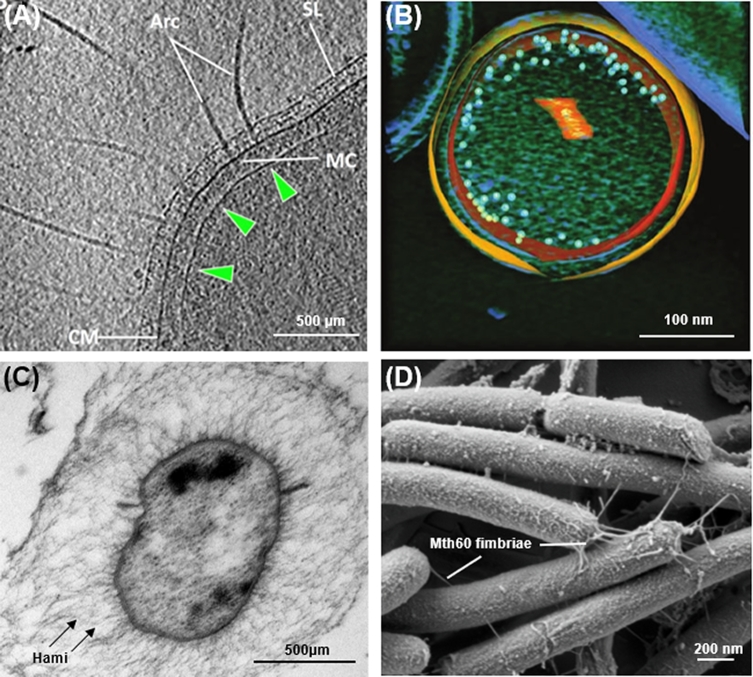
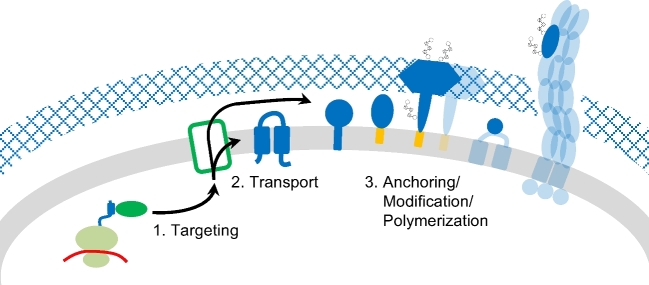
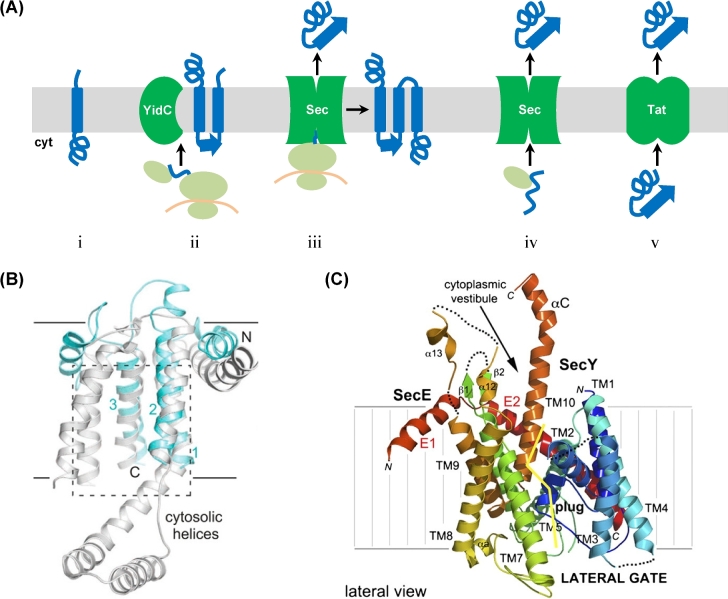
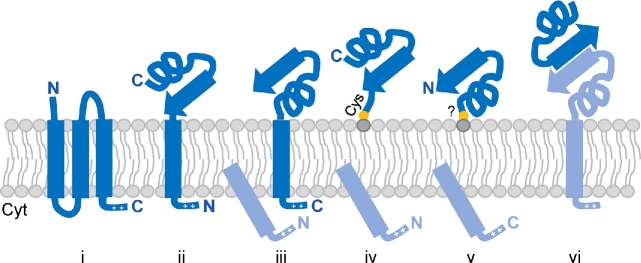
References
-
- Abdul Halim MF, Pfeiffer F, Zou J et al. Haloferax volcanii archaeosortase is required for motility, mating, and C-terminal processing of the S-layer glycoprotein. Mol Microbiol 2013;88:1164–75. - PubMed
-
- Abdul Halim MF, Rodriguez R, Stoltzfus JD et al. Conserved residues are critical for Haloferax volcanii archaeosortase catalytic activity: Implications for convergent evolution of the catalytic mechanisms of non-homologous sortases from archaea and bacteria. Mol Microbiol 2018;108:276–87. - PubMed
-
- Abu-Qarn M, Yurist-Doutsch S, Giordano A et al. Haloferax volcanii AglB and AglD are involved in N-glycosylation of the S-layer glycoprotein and proper assembly of the surface layer. J Mol Biol 2007;374:1224–36. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
