

RD5-mediated lack of PE_PGRS and PPE-MPTR export in BCG vaccine strains results in strong reduction of antigenic repertoire but little impact on protection
- PMID: 29912964
- PMCID: PMC6023246
- DOI: 10.1371/journal.ppat.1007139
RD5-mediated lack of PE_PGRS and PPE-MPTR export in BCG vaccine strains results in strong reduction of antigenic repertoire but little impact on protection
Abstract
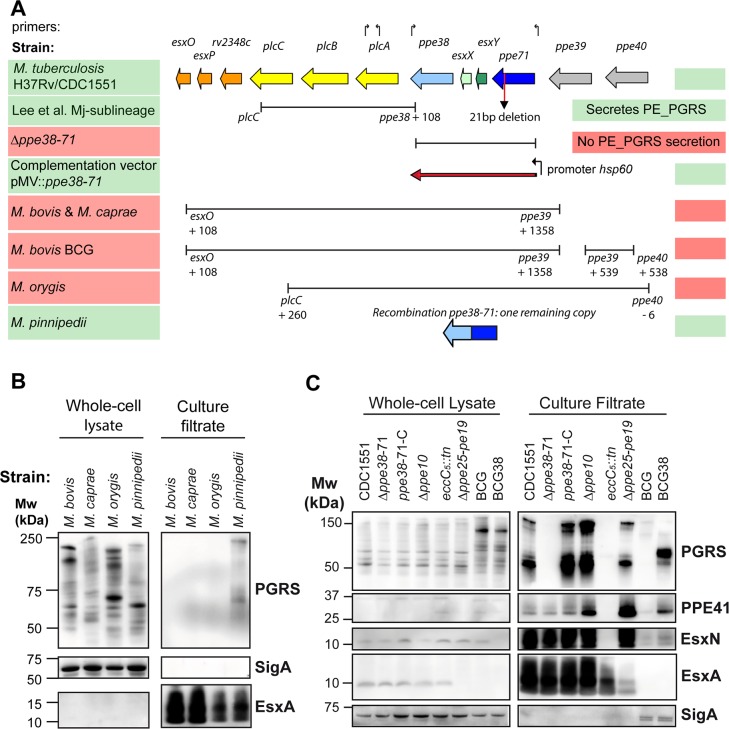
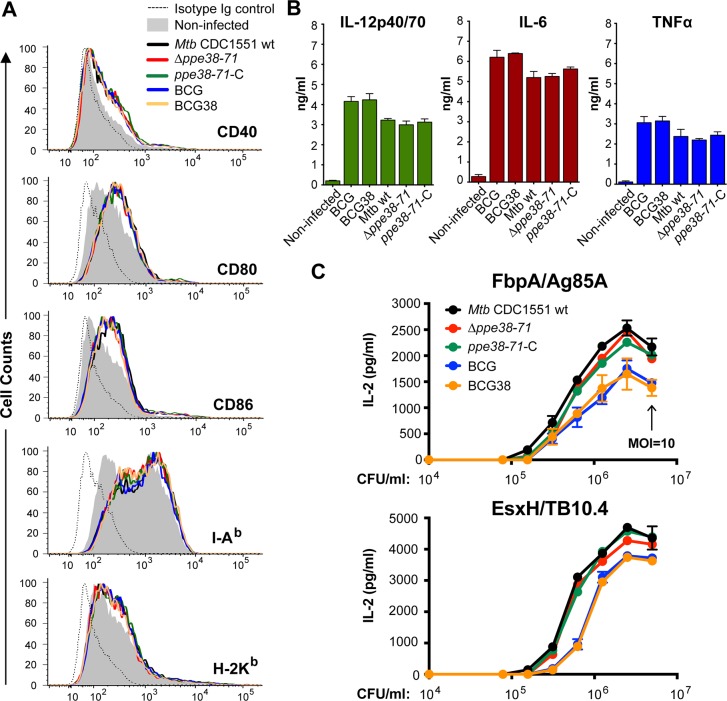
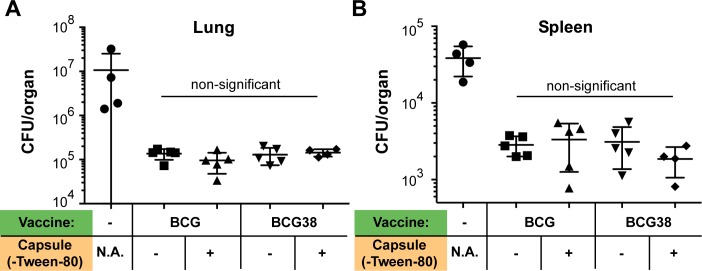
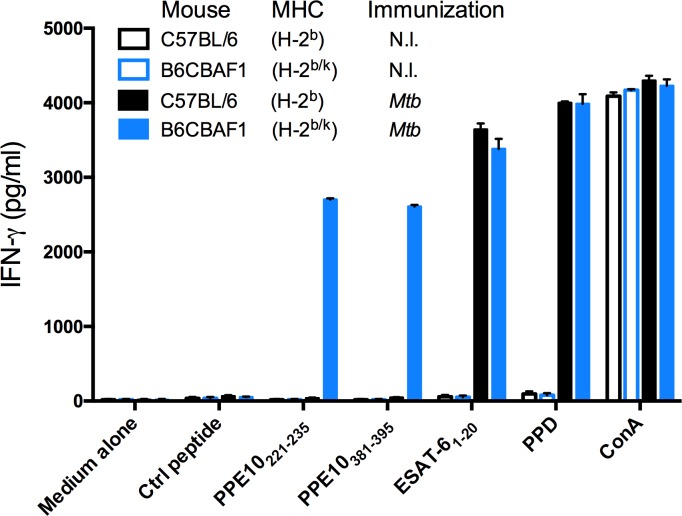
Tuberculosis is the deadliest infectious disease worldwide. Although the BCG vaccine is widely used, it does not efficiently protect against pulmonary tuberculosis and an improved tuberculosis vaccine is therefore urgently needed. Mycobacterium tuberculosis uses different ESX/Type VII secretion (T7S) systems to transport proteins important for virulence and host immune responses. We recently reported that secretion of T7S substrates belonging to the mycobacteria-specific Pro-Glu (PE) and Pro-Pro-Glu (PPE) proteins of the PGRS (polymorphic GC-rich sequences) and MPTR (major polymorphic tandem repeat) subfamilies required both a functional ESX-5 system and a functional PPE38/71 protein for secretion. Inactivation of ppe38/71 and the resulting loss of PE_PGRS/PPE-MPTR secretion were linked to increased virulence of M. tuberculosis strains. Here, we show that a predicted total of 89 PE_PGRS/PPE-MPTR surface proteins are not exported by certain animal-adapted strains of the M. tuberculosis complex including M. bovis. This Δppe38/71-associated secretion defect therefore also occurs in the M. bovis-derived tuberculosis vaccine BCG and could be partially restored by introduction of the M. tuberculosis ppe38-locus. Epitope mapping of the PPE-MPTR protein PPE10, further allowed us to monitor T-cell responses in splenocytes from BCG/M. tuberculosis immunized mice, confirming the dependence of PPE10-specific immune-induction on ESX-5/PPE38-mediated secretion. Restoration of PE_PGRS/PPE-MPTR secretion in recombinant BCG neither altered global antigenic presentation or activation of innate immune cells, nor protective efficacy in two different mouse vaccination-infection models. This unexpected finding stimulates a reassessment of the immunomodulatory properties of PE_PGRS/PPE-MPTR proteins, some of which are contained in vaccine formulations currently in clinical evaluation.
Conflict of interest statement
LM and RB are named inventors on a patent related to RD1, RD5 and RD8 regions of BCG. MAB is a named inventor on a separate patent related to genomic differences of the Mycobacterium tuberculosis complex. The other authors declare that no financial or competing interests exist.
Figures
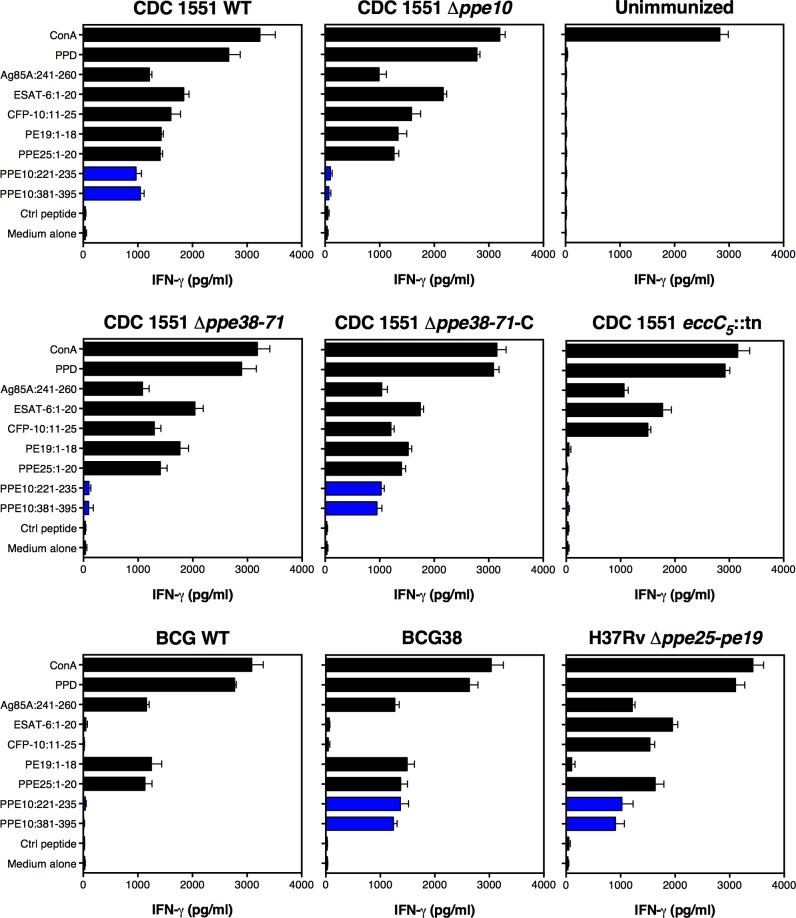
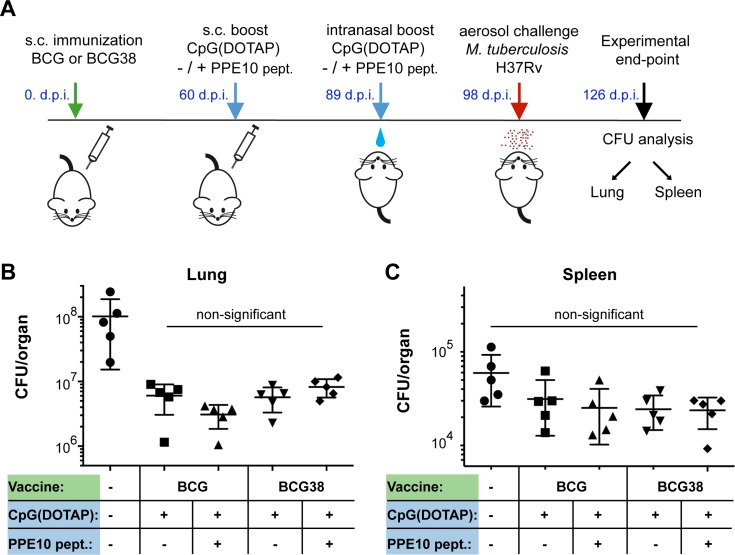
References
-
- WHO. Global tuberculosis report 2017. WHO. World Health Organization; 2017; Available: http://www.who.int/tb/publications/global_report/en/
-
- Arnold A, Witney AA, Vergnano S, Roche A, Cosgrove CA, Houston A, et al. XDR-TB transmission in London: Case management and contact tracing investigation assisted by early whole genome sequencing. J Infect. 2016;73(3):210–8. doi: 10.1016/j.jinf.2016.04.037 - DOI - PubMed
-
- Dheda K, Limberis JD, Pietersen E, Phelan J, Esmail A, Lesosky M, et al. Outcomes, infectiousness, and transmission dynamics of patients with extensively drug-resistant tuberculosis and home-discharged patients with programmatically incurable tuberculosis: a prospective cohort study. Lancet Respir Med. 2017;5(4):269–281. doi: 10.1016/S2213-2600(16)30433-7 - DOI - PubMed
-
- Fletcher HA, Schrager L. TB vaccine development and the End TB Strategy: importance and current status. Trans R Soc Trop Med Hyg. 2016;110: 212–218. doi: 10.1093/trstmh/trw016 - DOI - PMC - PubMed
-
- Behr M., Small P. A historical and molecular phylogeny of BCG strains. Vaccine. 1999;17: 915–922. doi: 10.1016/S0264-410X(98)00277-1 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
