

Serine metabolism in the brain regulates starvation-induced sleep suppression in Drosophila melanogaster
- PMID: 29915051
- PMCID: PMC6142195
- DOI: 10.1073/pnas.1719033115
Serine metabolism in the brain regulates starvation-induced sleep suppression in Drosophila melanogaster
Abstract
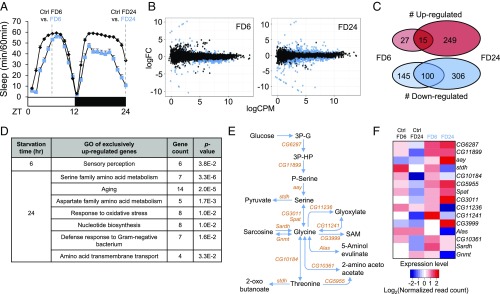
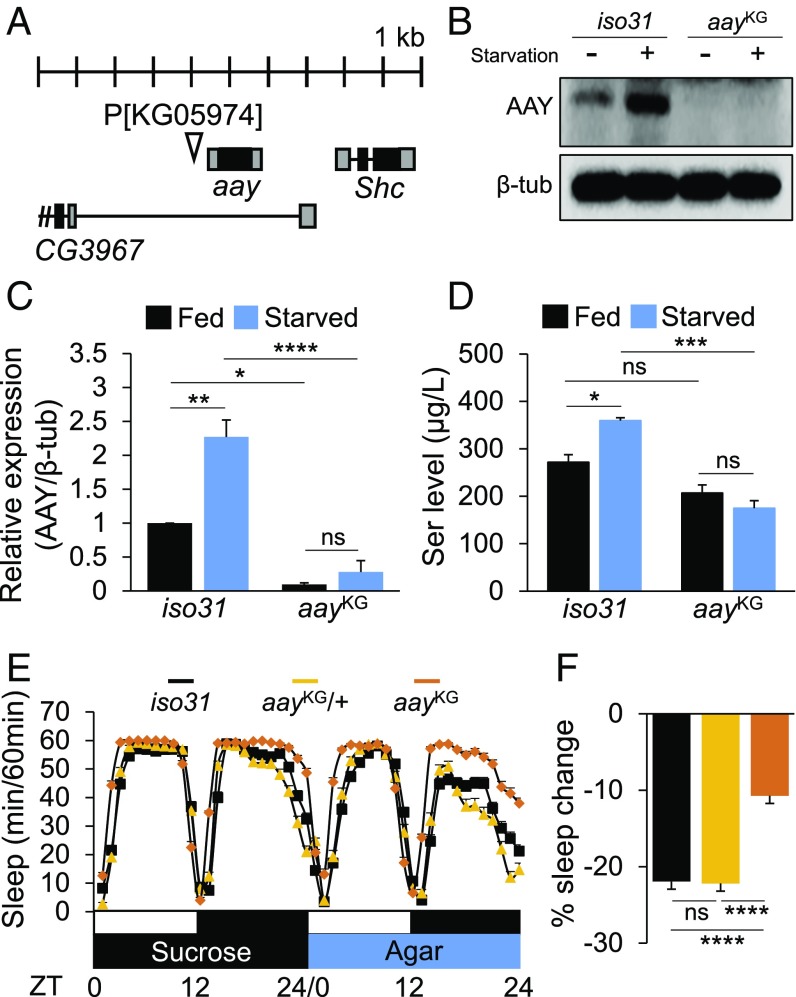
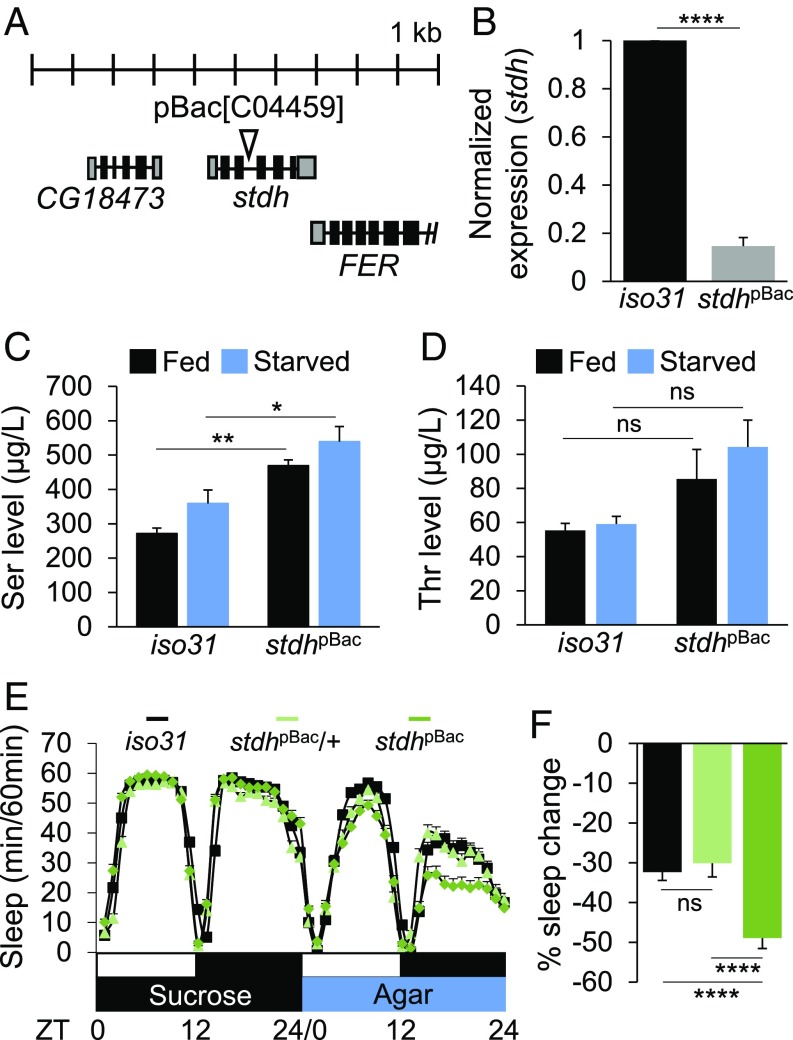
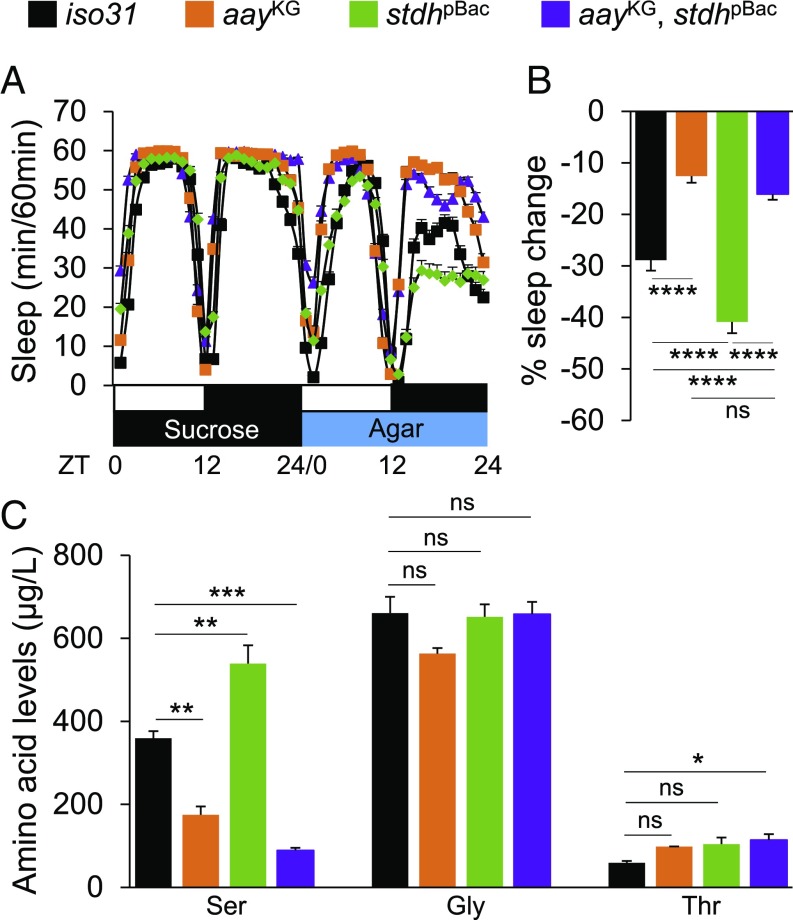
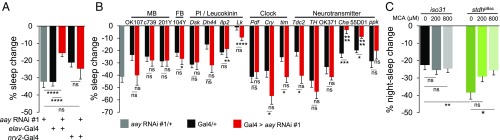
Sleep and metabolism are physiologically and behaviorally intertwined; however, the molecular basis for their interaction remains poorly understood. Here, we identified a serine metabolic pathway as a key mediator for starvation-induced sleep suppression. Transcriptome analyses revealed that enzymes involved in serine biosynthesis were induced upon starvation in Drosophila melanogaster brains. Genetic mutants of astray (aay), a fly homolog of the rate-limiting phosphoserine phosphatase in serine biosynthesis, displayed reduced starvation-induced sleep suppression. In contrast, a hypomorphic mutation in a serine/threonine-metabolizing enzyme, serine/threonine dehydratase (stdh), exaggerated starvation-induced sleep suppression. Analyses of double mutants indicated that aay and stdh act on the same genetic pathway to titrate serine levels in the head as well as to adjust starvation-induced sleep behaviors. RNA interference-mediated depletion of aay expression in neurons, using cholinergic Gal4 drivers, phenocopied aay mutants, while a nicotinic acetylcholine receptor antagonist selectively rescued the exaggerated starvation-induced sleep suppression in stdh mutants. Taken together, these data demonstrate that neural serine metabolism controls sleep during starvation, possibly via cholinergic signaling. We propose that animals have evolved a sleep-regulatory mechanism that reprograms amino acid metabolism for adaptive sleep behaviors in response to metabolic needs.
Keywords: serine; sleep regulation; starvation.
Copyright © 2018 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Spiegel K, Tasali E, Penev P, Van Cauter E. Brief communication: Sleep curtailment in healthy young men is associated with decreased leptin levels, elevated ghrelin levels, and increased hunger and appetite. Ann Intern Med. 2004;141:846–850. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
