

Molecular determination of claudin-15 organization and channel selectivity
- PMID: 29915162
- PMCID: PMC6028499
- DOI: 10.1085/jgp.201711868
Molecular determination of claudin-15 organization and channel selectivity
Abstract
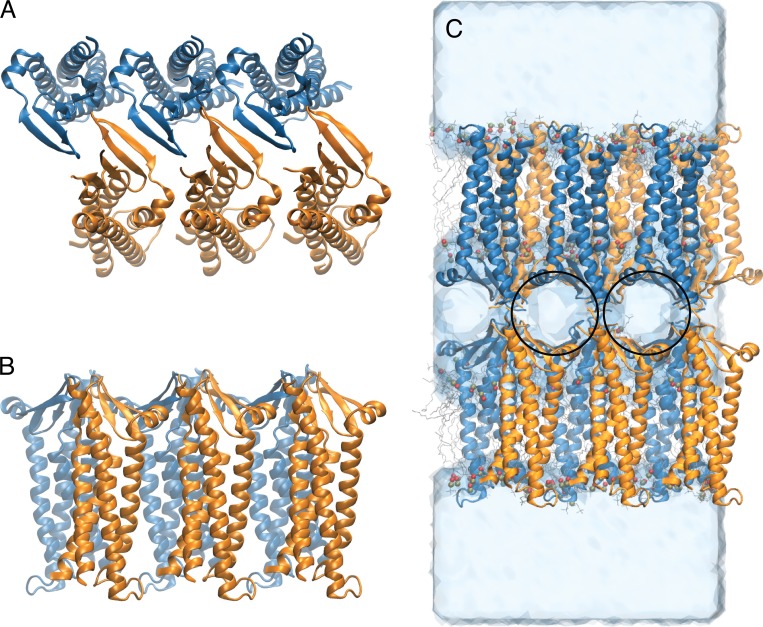
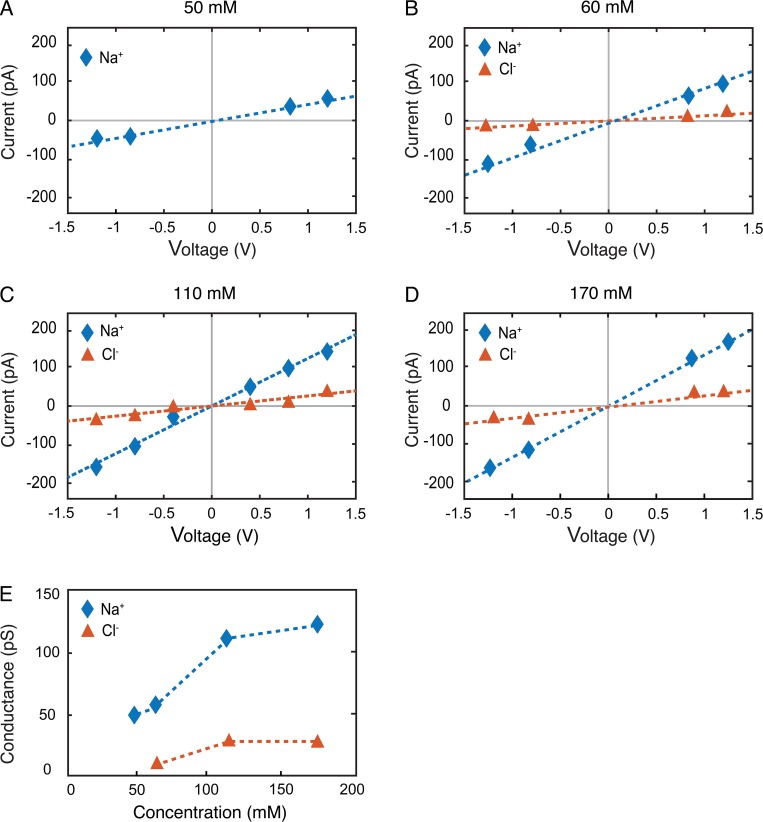
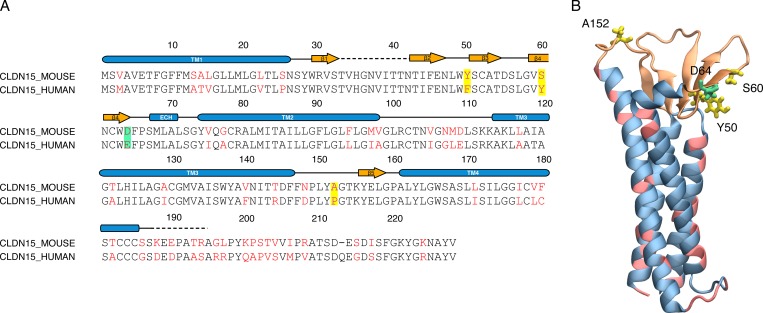
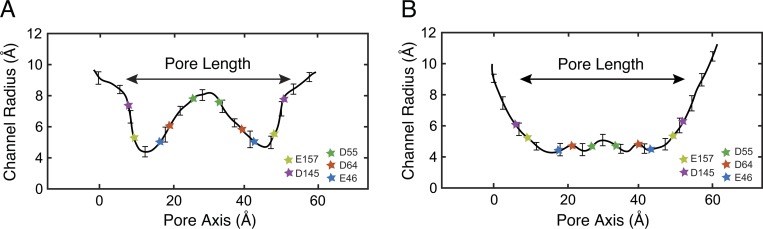
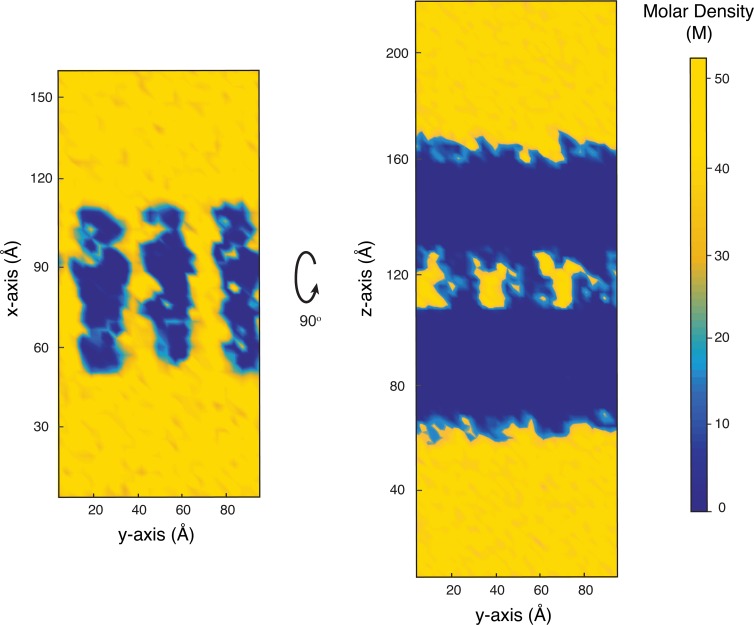
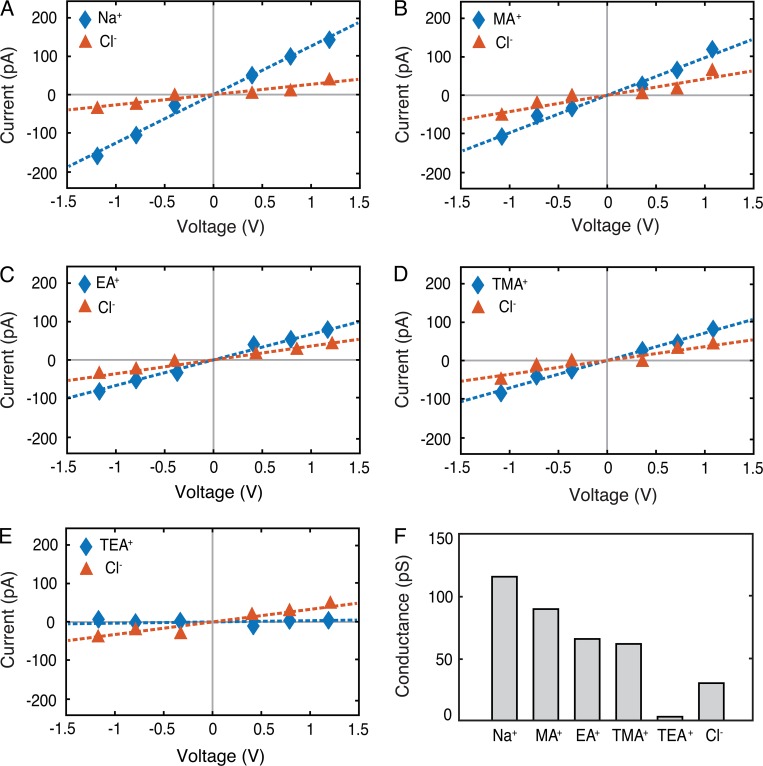
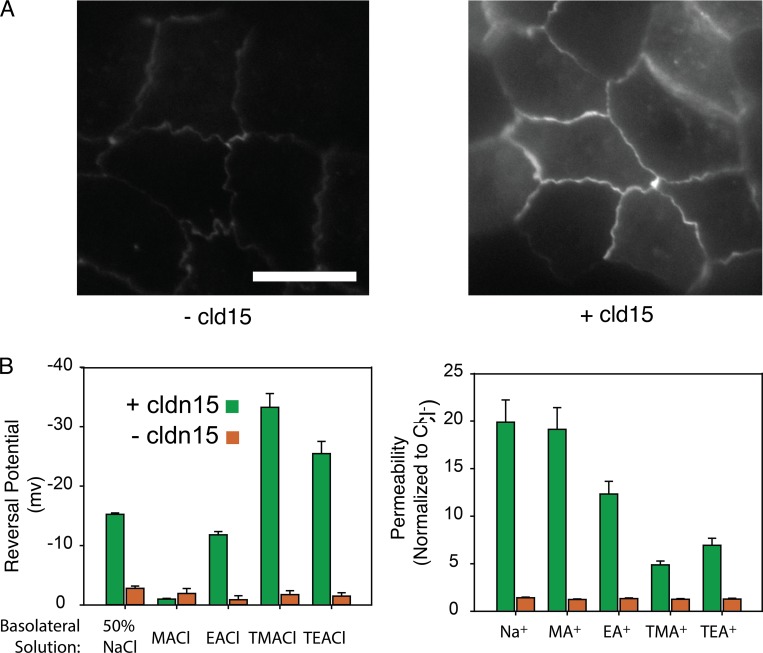
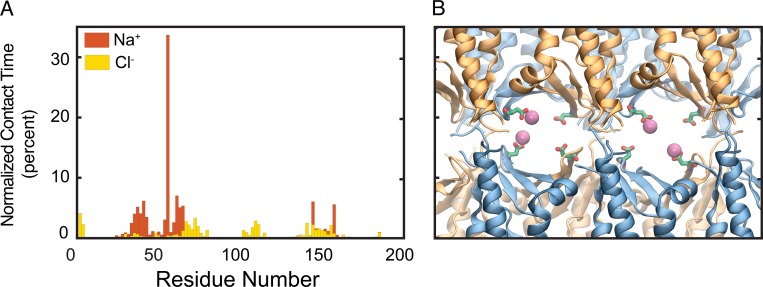
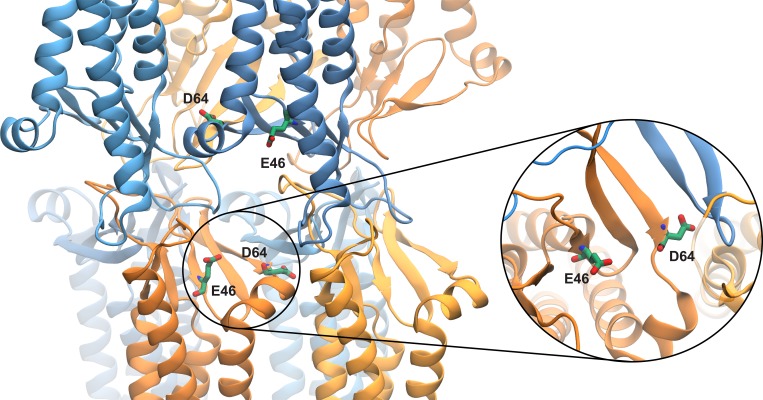
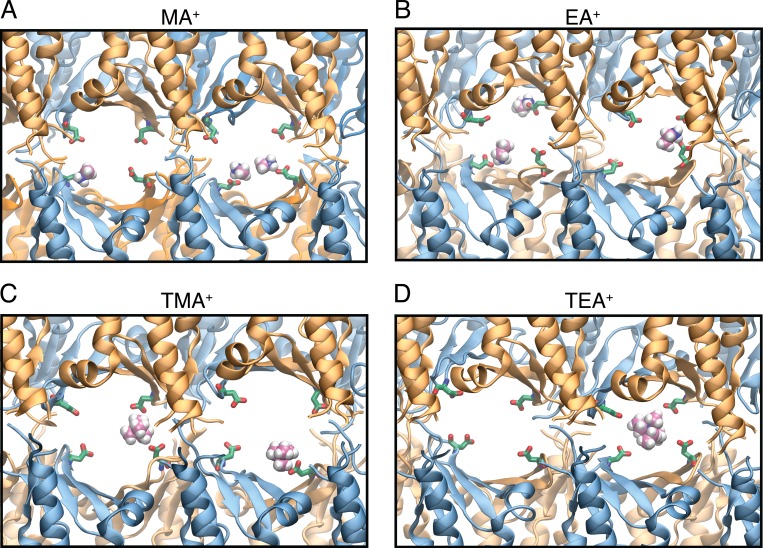
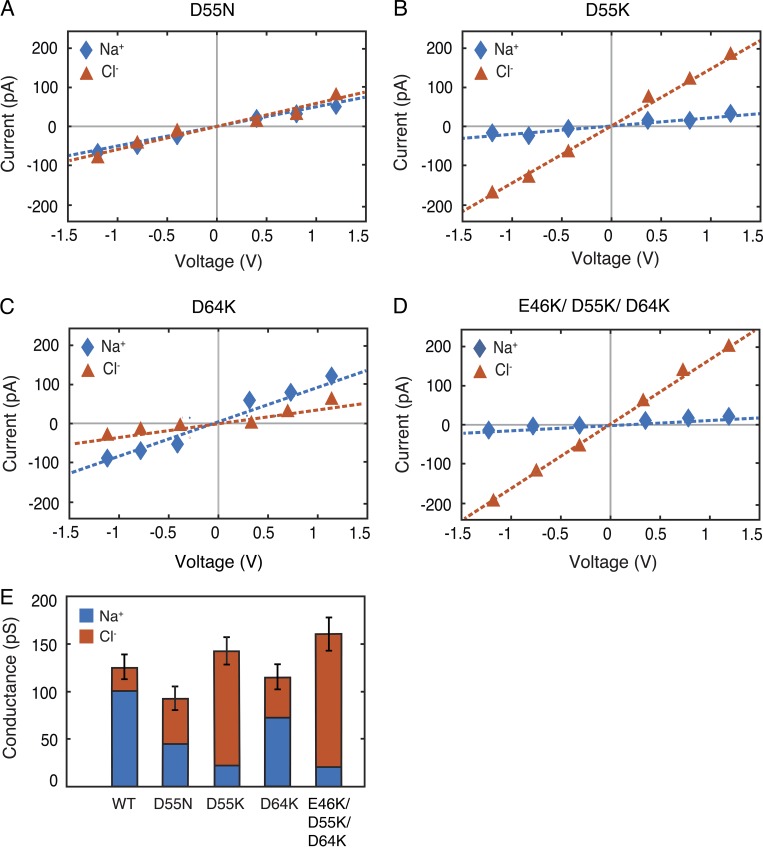
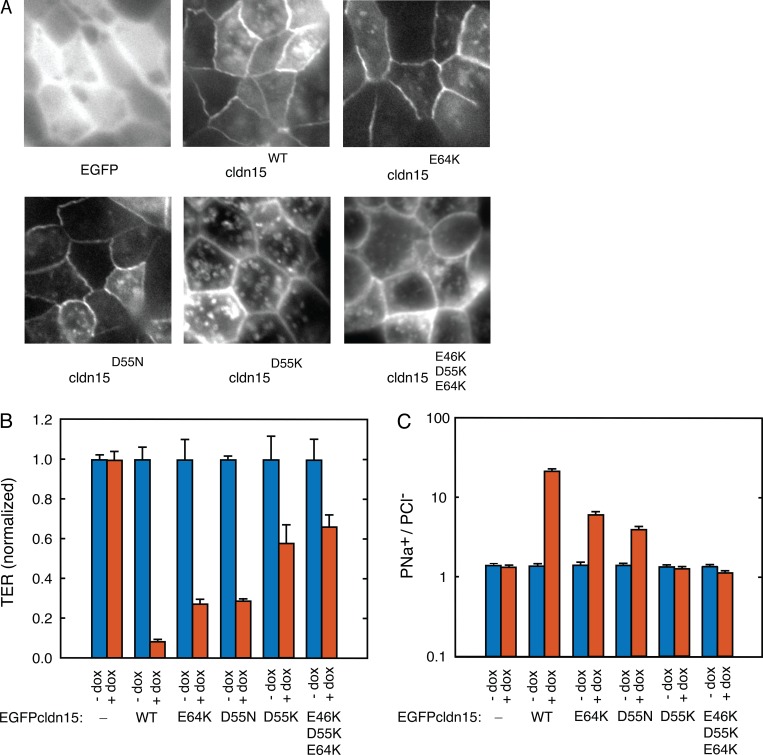
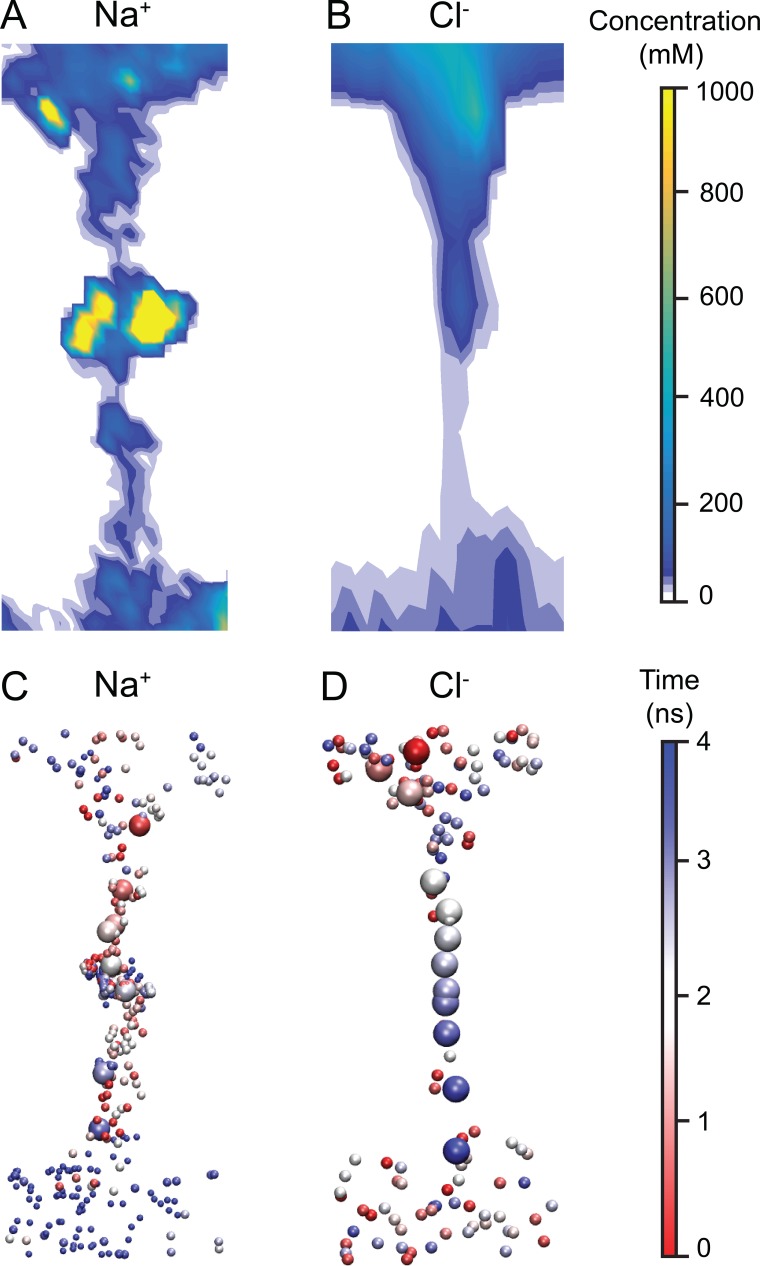
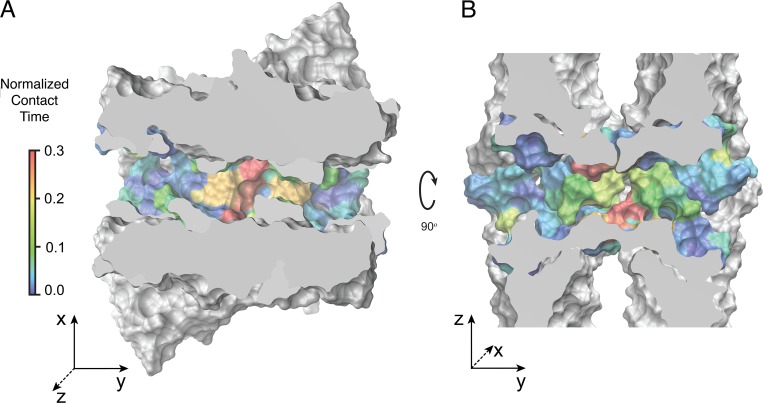
Tight junctions are macromolecular structures that traverse the space between adjacent cells in epithelia and endothelia. Members of the claudin family are known to determine tight junction permeability in a charge- and size-selective manner. Here, we use molecular dynamics simulations to build and refine an atomic model of claudin-15 channels and study its transport properties. Our simulations indicate that claudin-15 forms well-defined channels for ions and molecules and otherwise "seals" the paracellular space through hydrophobic interactions. Ionic currents, calculated from simulation trajectories of wild-type as well as mutant channels, reflect in vitro measurements. The simulations suggest that the selectivity filter is formed by a cage of four aspartic acid residues (D55), contributed by four claudin-15 molecules, which creates a negative electrostatic potential to favor cation flux over anion flux. Charge reversal or charge ablation mutations of D55 significantly reduce cation permeability in silico and in vitro, whereas mutations of other negatively charged pore amino acid residues have a significantly smaller impact on channel permeability and selectivity. The simulations also indicate that water and small ions can pass through the channel, but larger cations, such as tetramethylammonium, do not traverse the pore. Thus, our model provides an atomic view of claudin channels, their transport function, and a potential three-dimensional organization of its selectivity filter.
© 2018 Khalili-Araghi et al.
Figures
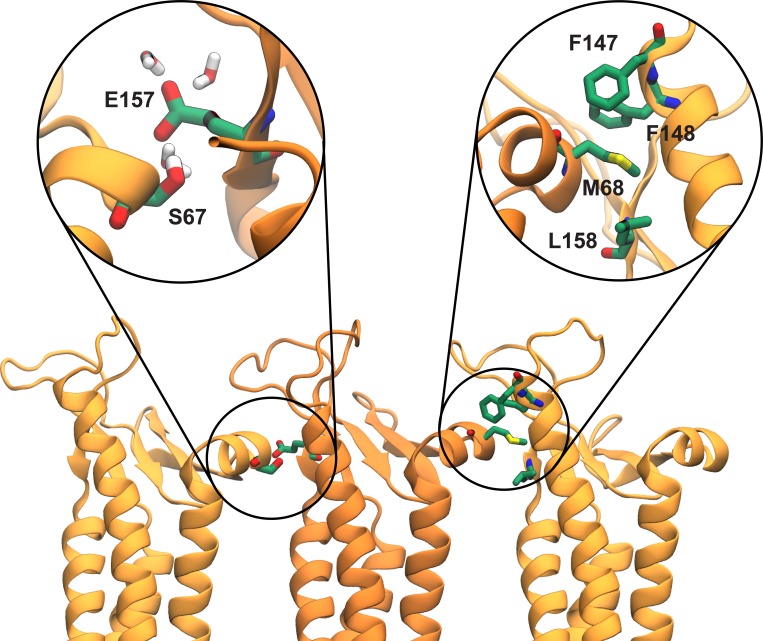
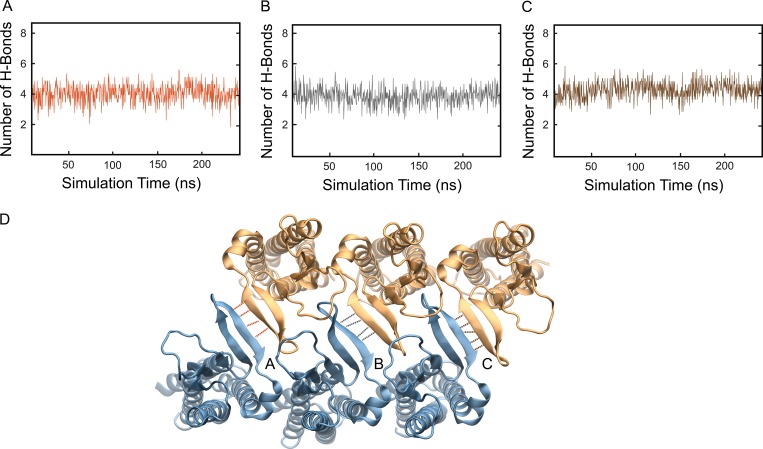
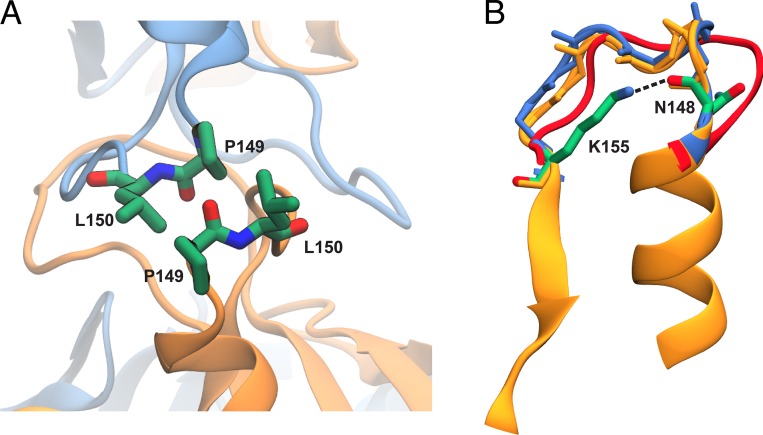
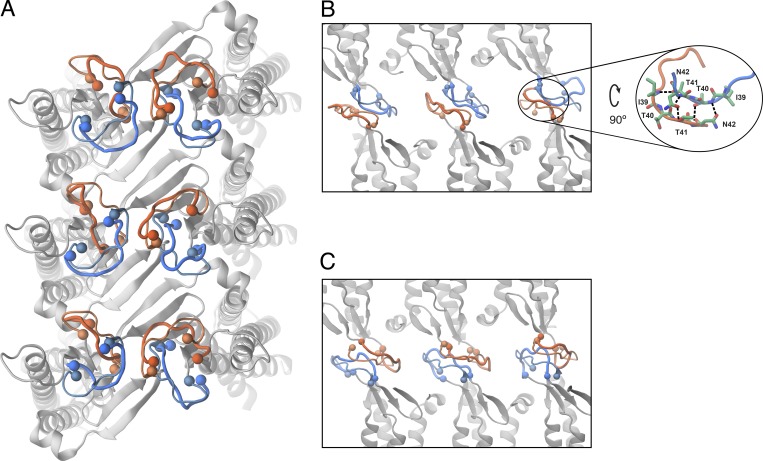
Comment in
-
Claudins get a closer look.J Gen Physiol. 2018 Jul 2;150(7):893. doi: 10.1085/jgp.201812136. Epub 2018 Jun 19. J Gen Physiol. 2018. PMID: 29921660 Free PMC article.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
