

A CD4-mimetic compound enhances vaccine efficacy against stringent immunodeficiency virus challenge
- PMID: 29915222
- PMCID: PMC6006336
- DOI: 10.1038/s41467-018-04758-9
A CD4-mimetic compound enhances vaccine efficacy against stringent immunodeficiency virus challenge
Abstract
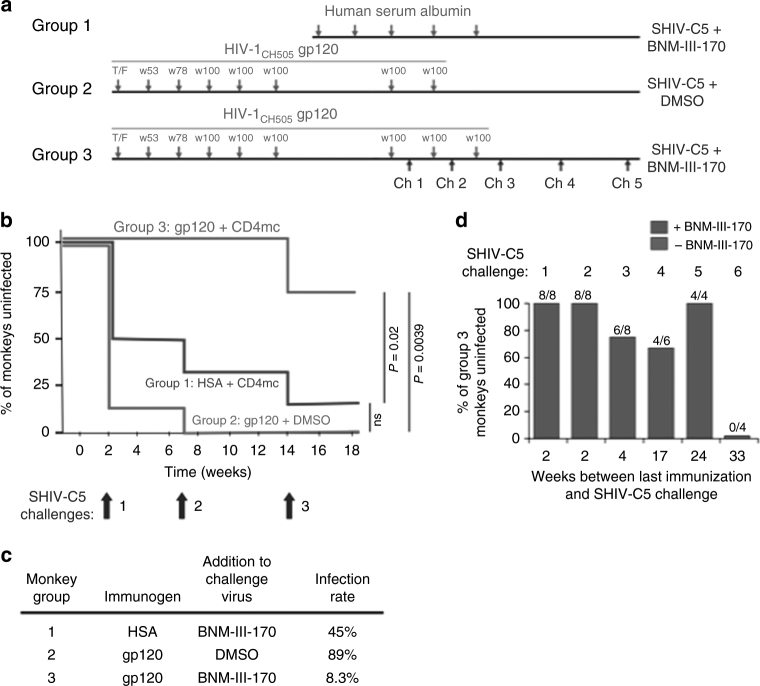
The envelope glycoprotein (Env) trimer ((gp120/gp41)3) mediates human immunodeficiency virus (HIV-1) entry into cells. The "closed," antibody-resistant Env trimer is driven to more open conformations by binding the host receptor, CD4. Broadly neutralizing antibodies that recognize conserved elements of the closed Env are potentially protective, but are elicited inefficiently. HIV-1 has evolved multiple mechanisms to evade readily elicited antibodies against more open Env conformations. Small-molecule CD4-mimetic compounds (CD4mc) bind the HIV-1 gp120 Env and promote conformational changes similar to those induced by CD4, exposing conserved Env elements to antibodies. Here, we show that a CD4mc synergizes with antibodies elicited by monomeric HIV-1 gp120 to protect monkeys from multiple high-dose intrarectal challenges with a heterologous simian-human immunodeficiency virus (SHIV). The protective immune response persists for at least six months after vaccination. CD4mc should increase the protective efficacy of any HIV-1 Env vaccine that elicits antibodies against CD4-induced conformations of Env.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Characterization of Human Immunodeficiency Virus (HIV-1) Envelope Glycoprotein Variants Selected for Resistance to a CD4-Mimetic Compound.J Virol. 2022 Sep 14;96(17):e0063622. doi: 10.1128/jvi.00636-22. Epub 2022 Aug 18. J Virol. 2022. PMID: 35980207 Free PMC article.
-
Activation and Inactivation of Primary Human Immunodeficiency Virus Envelope Glycoprotein Trimers by CD4-Mimetic Compounds.J Virol. 2017 Jan 18;91(3):e01880-16. doi: 10.1128/JVI.01880-16. Print 2017 Feb 1. J Virol. 2017. PMID: 27881646 Free PMC article.
-
Cross-Linking of a CD4-Mimetic Miniprotein with HIV-1 Env gp140 Alters Kinetics and Specificities of Antibody Responses against HIV-1 Env in Macaques.J Virol. 2017 Sep 12;91(19):e00401-17. doi: 10.1128/JVI.00401-17. Print 2017 Oct 1. J Virol. 2017. PMID: 28490585 Free PMC article.
-
Vaccination of rhesus macaques with recombinant Mycobacterium bovis bacillus Calmette-Guérin Env V3 elicits neutralizing antibody-mediated protection against simian-human immunodeficiency virus with a homologous but not a heterologous V3 motif.J Virol. 2005 Feb;79(3):1452-62. doi: 10.1128/JVI.79.3.1452-1462.2005. J Virol. 2005. PMID: 15650171 Free PMC article.
-
An HIV Envelope gp120-Fc Fusion Protein Elicits Effector Antibody Responses in Rhesus Macaques.Clin Vaccine Immunol. 2017 Jun 5;24(6):e00028-17. doi: 10.1128/CVI.00028-17. Print 2017 Jun. Clin Vaccine Immunol. 2017. PMID: 28404572 Free PMC article.
Cited by
-
HIV-1 Envelope Glycoproteins Proteolytic Cleavage Protects Infected Cells from ADCC Mediated by Plasma from Infected Individuals.Viruses. 2021 Nov 6;13(11):2236. doi: 10.3390/v13112236. Viruses. 2021. PMID: 34835042 Free PMC article.
-
The Conformational States of the HIV-1 Envelope Glycoproteins.Trends Microbiol. 2020 Aug;28(8):655-667. doi: 10.1016/j.tim.2020.03.007. Epub 2020 May 14. Trends Microbiol. 2020. PMID: 32418859 Free PMC article. Review.
-
Indoline CD4-mimetic compounds mediate potent and broad HIV-1 inhibition and sensitization to antibody-dependent cellular cytotoxicity.Proc Natl Acad Sci U S A. 2023 Mar 28;120(13):e2222073120. doi: 10.1073/pnas.2222073120. Epub 2023 Mar 24. Proc Natl Acad Sci U S A. 2023. PMID: 36961924 Free PMC article.
-
A New Family of Small-Molecule CD4-Mimetic Compounds Contacts Highly Conserved Aspartic Acid 368 of HIV-1 gp120 and Mediates Antibody-Dependent Cellular Cytotoxicity.J Virol. 2019 Nov 26;93(24):e01325-19. doi: 10.1128/JVI.01325-19. Print 2019 Dec 15. J Virol. 2019. PMID: 31554684 Free PMC article.
-
SERINC5 Restricts HIV-1 Infectivity by Promoting Conformational Changes and Accelerating Functional Inactivation of Env.Viruses. 2022 Jun 25;14(7):1388. doi: 10.3390/v14071388. Viruses. 2022. PMID: 35891369 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
