

Scavenging of superoxide by a membrane-bound superoxide oxidase
- PMID: 29915379
- PMCID: PMC6379055
- DOI: 10.1038/s41589-018-0072-x
Scavenging of superoxide by a membrane-bound superoxide oxidase
Abstract
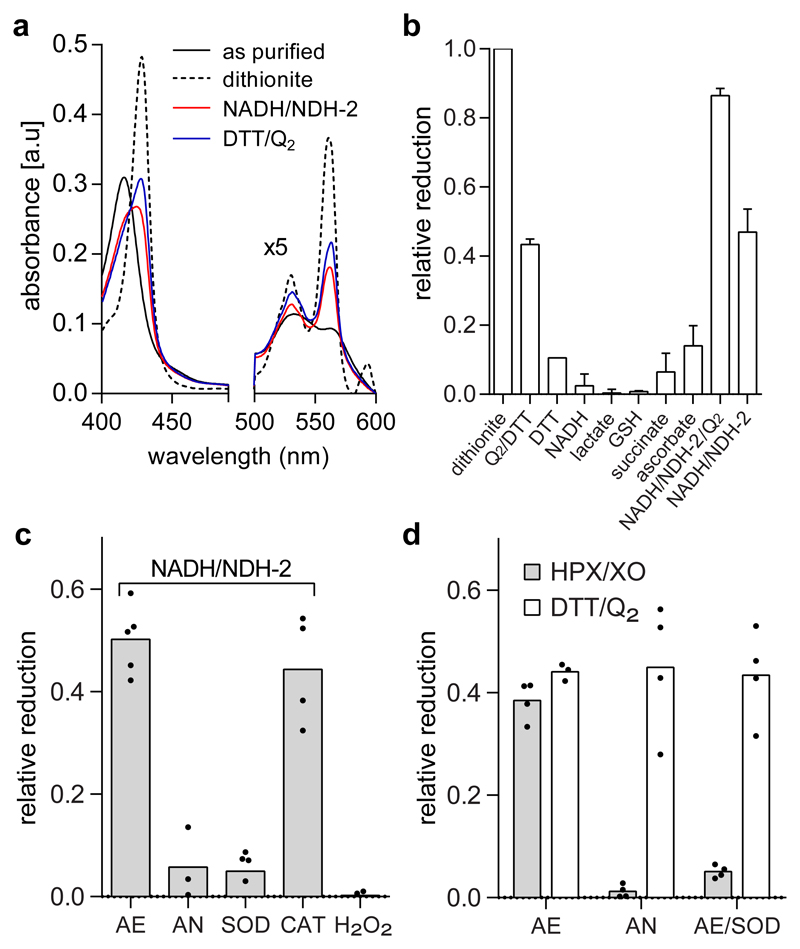
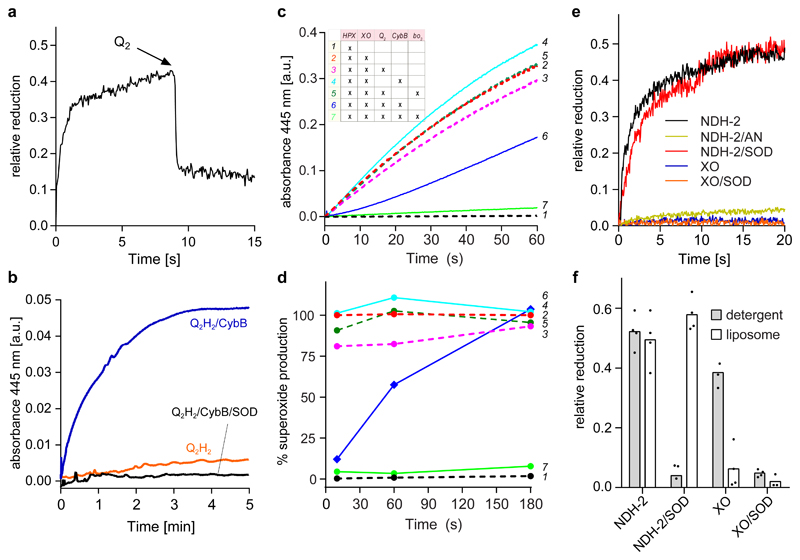
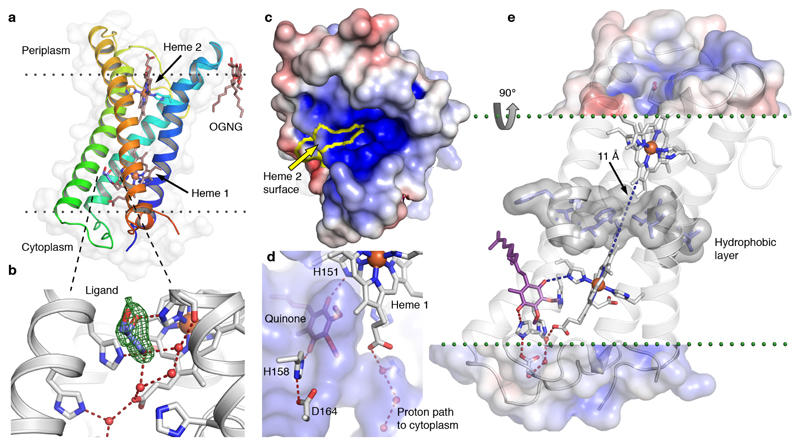
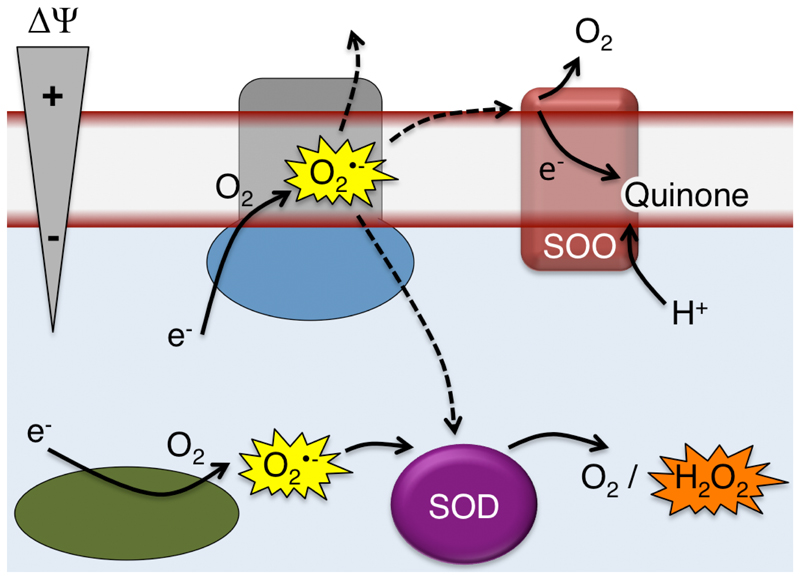
Superoxide is a reactive oxygen species produced during aerobic metabolism in mitochondria and prokaryotes. It causes damage to lipids, proteins and DNA and is implicated in cancer, cardiovascular disease, neurodegenerative disorders and aging. As protection, cells express soluble superoxide dismutases, disproportionating superoxide to oxygen and hydrogen peroxide. Here, we describe a membrane-bound enzyme that directly oxidizes superoxide and funnels the sequestered electrons to ubiquinone in a diffusion-limited reaction. Experiments in proteoliposomes and inverted membranes show that the protein is capable of efficiently quenching superoxide generated at the membrane in vitro. The 2.0 Å crystal structure shows an integral membrane di-heme cytochrome b poised for electron transfer from the P-side and proton uptake from the N-side. This suggests that the reaction is electrogenic and contributes to the membrane potential while also conserving energy by reducing the quinone pool. Based on this enzymatic activity, we propose that the enzyme family be denoted superoxide oxidase (SOO).
Conflict of interest statement
Figures
References
-
- Dröge W. Free radicals in the physiological control of cell function. Physiol Rev. 2002;82:47–95. - PubMed
-
- Barnham KJ, Masters CL, Bush AI. Neurodegenerative diseases and oxidative stress. Nat Rev Drug Discov. 2004;3:205–214. - PubMed
-
- Riera CE, Merkwirth C, De Magalhaes Filho CD, Dillin A. Signaling Networks Determining Life Span. Annu Rev Biochem. 2016;85:35–64. - PubMed
-
- McCord JM, Fridovich I. Superoxide dismutase. An enzymic function for erythrocuprein (hemocuprein) J Biol Chem. 1969;244:6049–6055. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
