

Virulence Associated Gene 8 of Bordetella pertussis Enhances Contact System Activity by Inhibiting the Regulatory Function of Complement Regulator C1 Inhibitor
- PMID: 29915576
- PMCID: PMC5994690
- DOI: 10.3389/fimmu.2018.01172
Virulence Associated Gene 8 of Bordetella pertussis Enhances Contact System Activity by Inhibiting the Regulatory Function of Complement Regulator C1 Inhibitor
Abstract
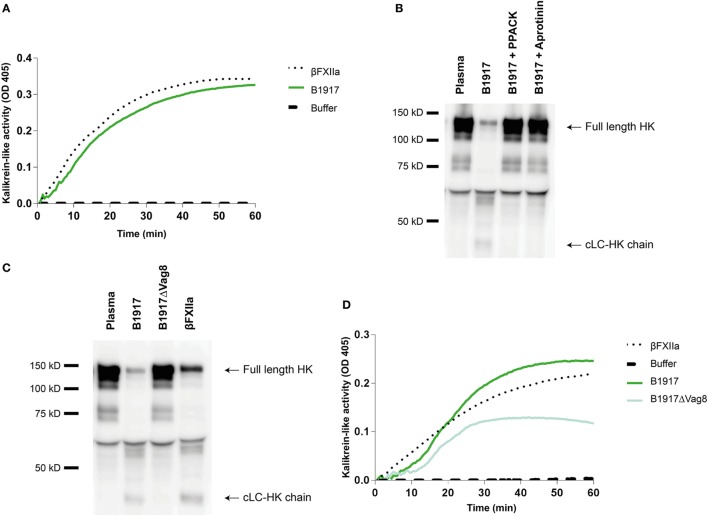
Bordetella pertussis is a Gram-negative bacterium and the causative agent of whooping cough. Whooping cough is currently re-emerging worldwide and, therefore, still poses a continuous global health threat. B. pertussis expresses several virulence factors that play a role in evading the human immune response. One of these virulence factors is virulence associated gene 8 (Vag8). Vag8 is a complement evasion molecule that mediates its effects by binding to the complement regulator C1 inhibitor (C1-INH). This regulatory protein is a fluid phase serine protease that controls proenzyme activation and enzyme activity of not only the complement system but also the contact system. Activation of the contact system results in the generation of bradykinin, a pro-inflammatory peptide. Here, the activation of the contact system by B. pertussis was explored. We demonstrate that recombinant as well as endogenous Vag8 enhanced contact system activity by binding C1-INH and attenuating its inhibitory function. Moreover, we show that B. pertussis itself is able to activate the contact system. This activation was dependent on Vag8 production as a Vag8 knockout B. pertussis strain was unable to activate the contact system. These findings show a previously overlooked interaction between the contact system and the respiratory pathogen B. pertussis. Activation of the contact system by B. pertussis may contribute to its pathogenicity and virulence.
Keywords: Bordetella pertussis; C1 inhibitor; contact system; virulence associated gene 8; whooping cough.
Figures
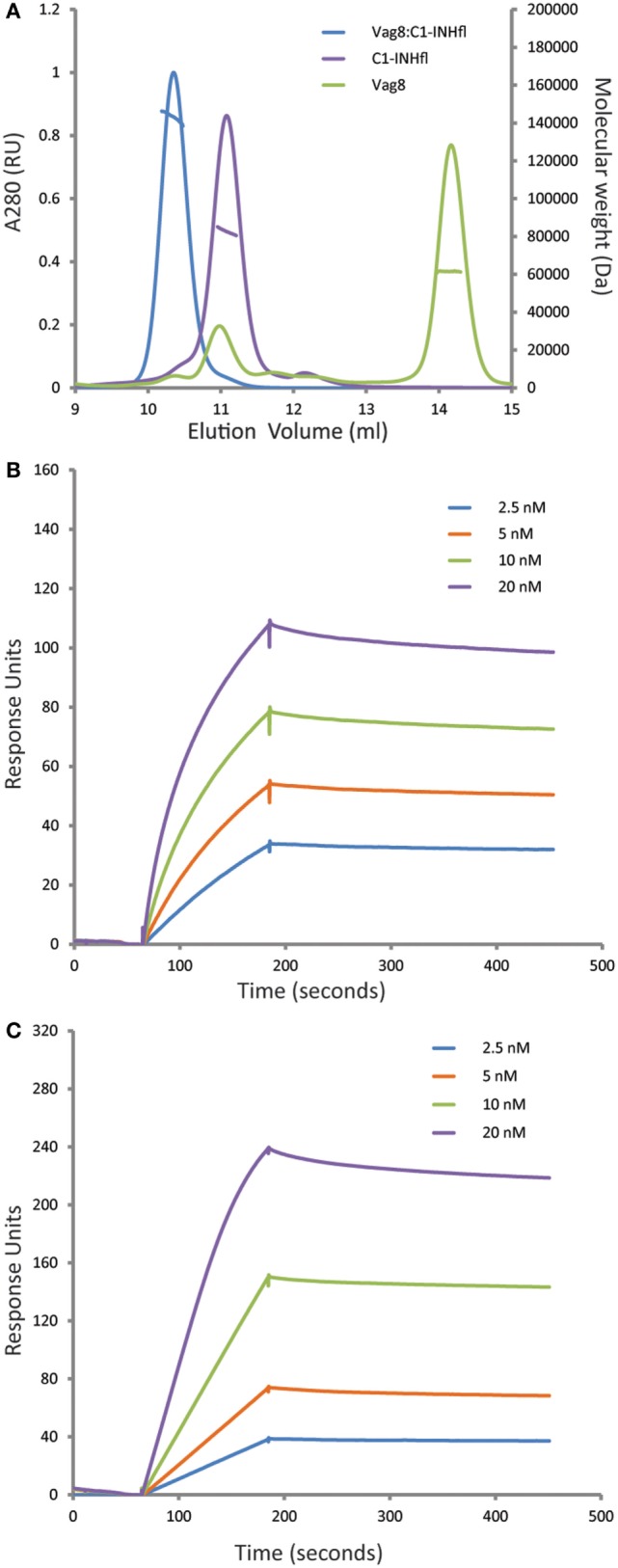
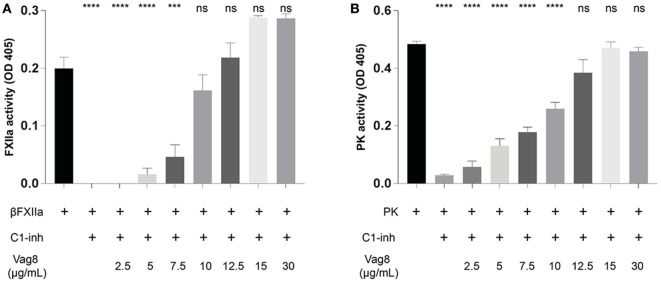
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous