

Itaconic acid mediates crosstalk between macrophage metabolism and peritoneal tumors
- PMID: 29920191
- PMCID: PMC6118601
- DOI: 10.1172/JCI99169
Itaconic acid mediates crosstalk between macrophage metabolism and peritoneal tumors
Abstract
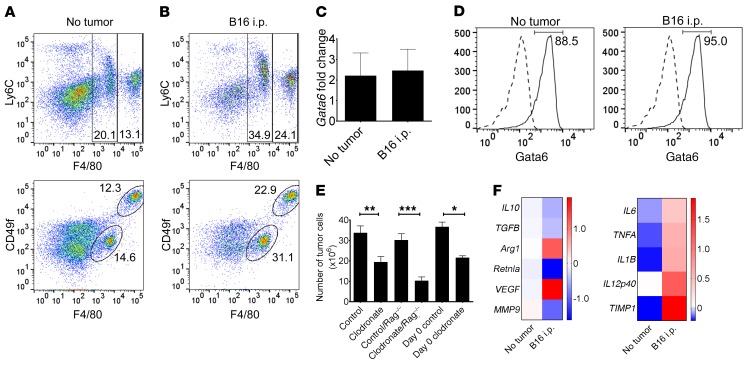
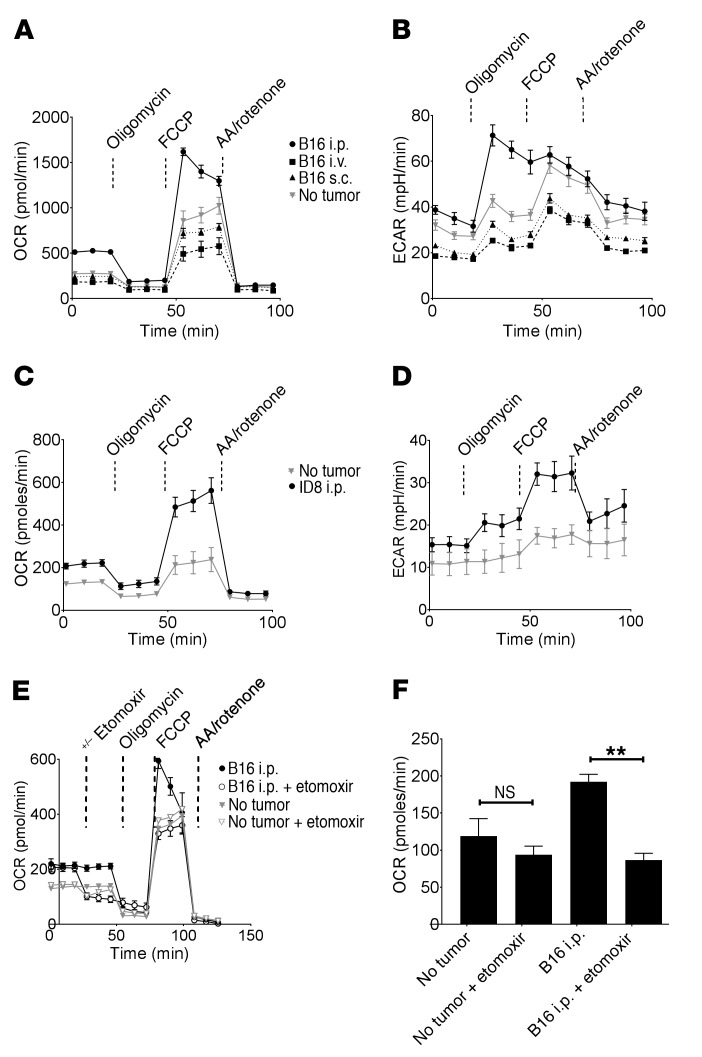
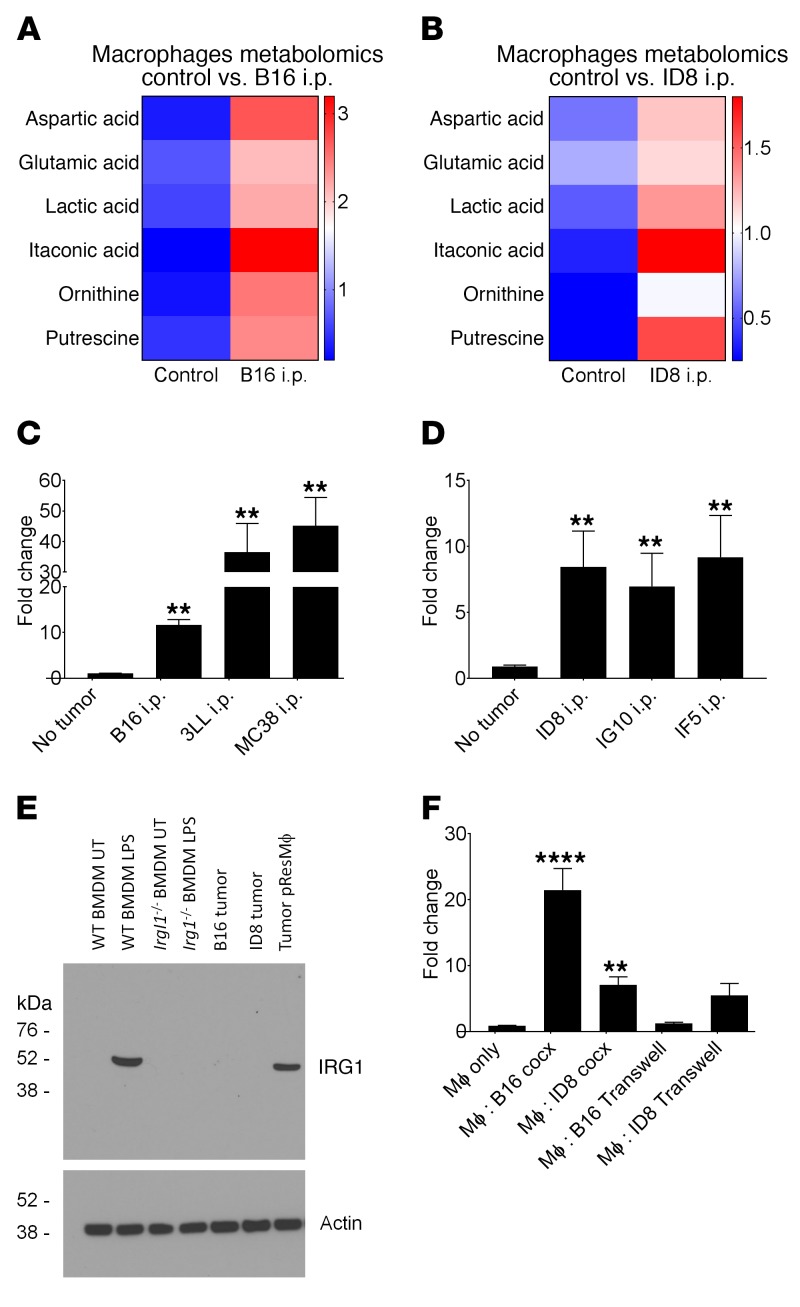
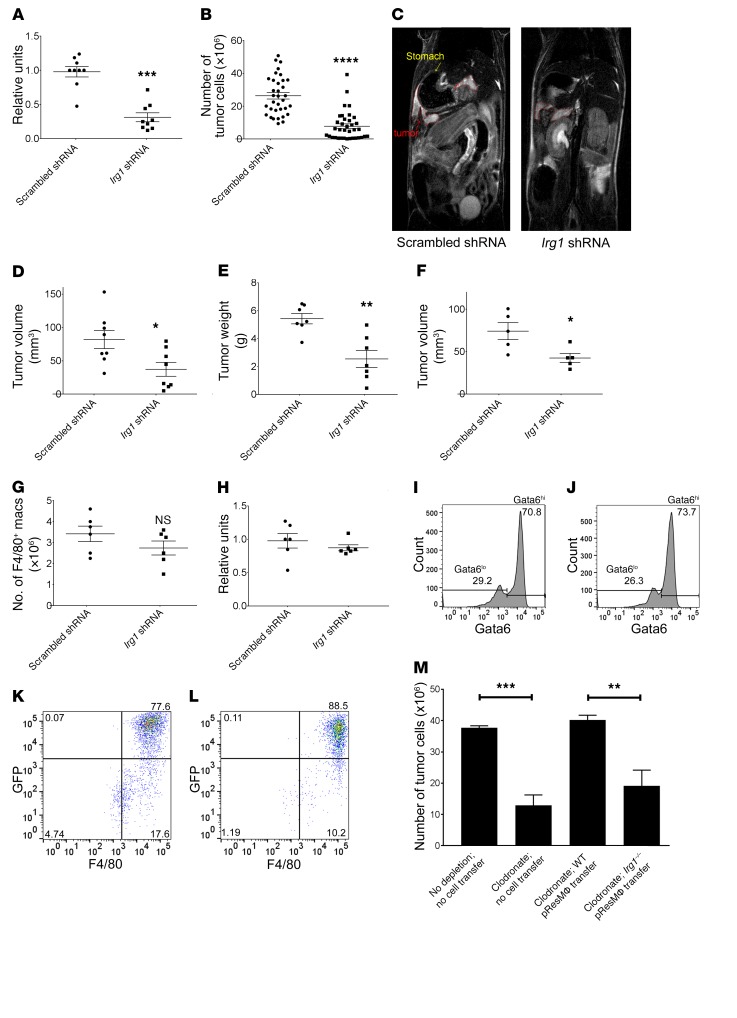
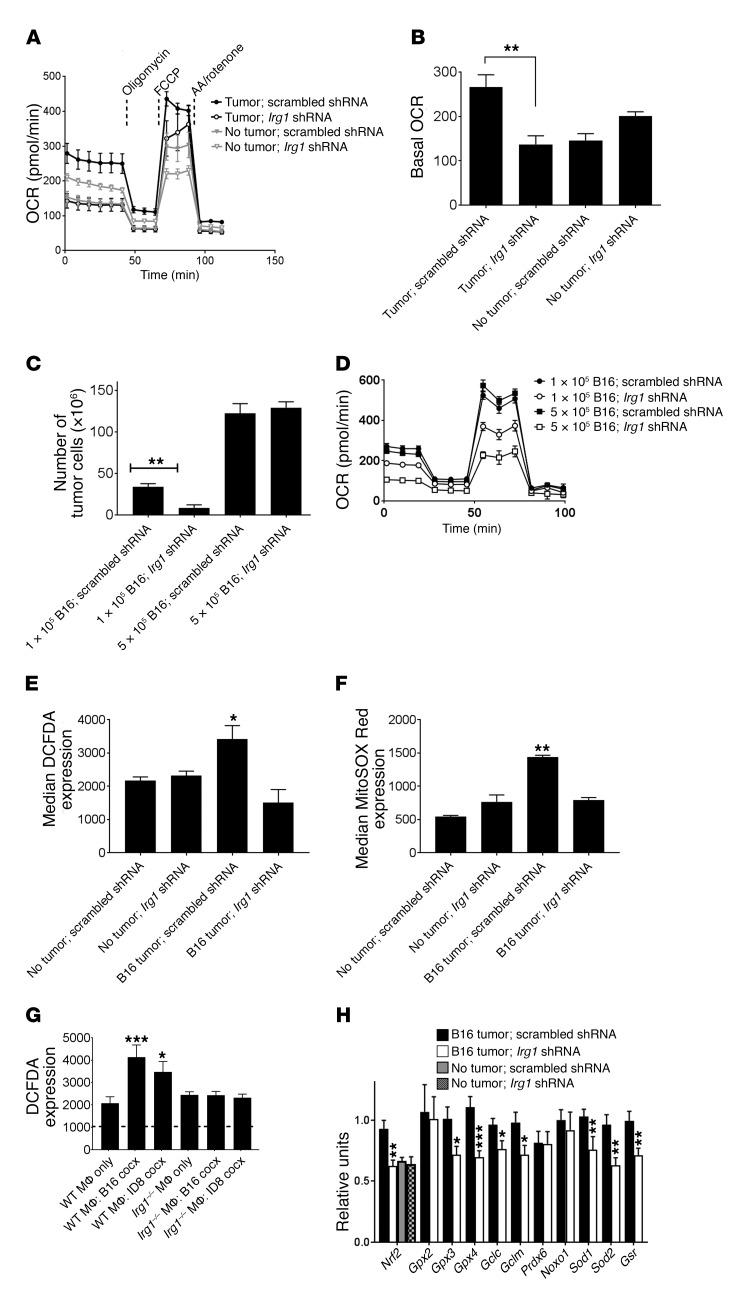
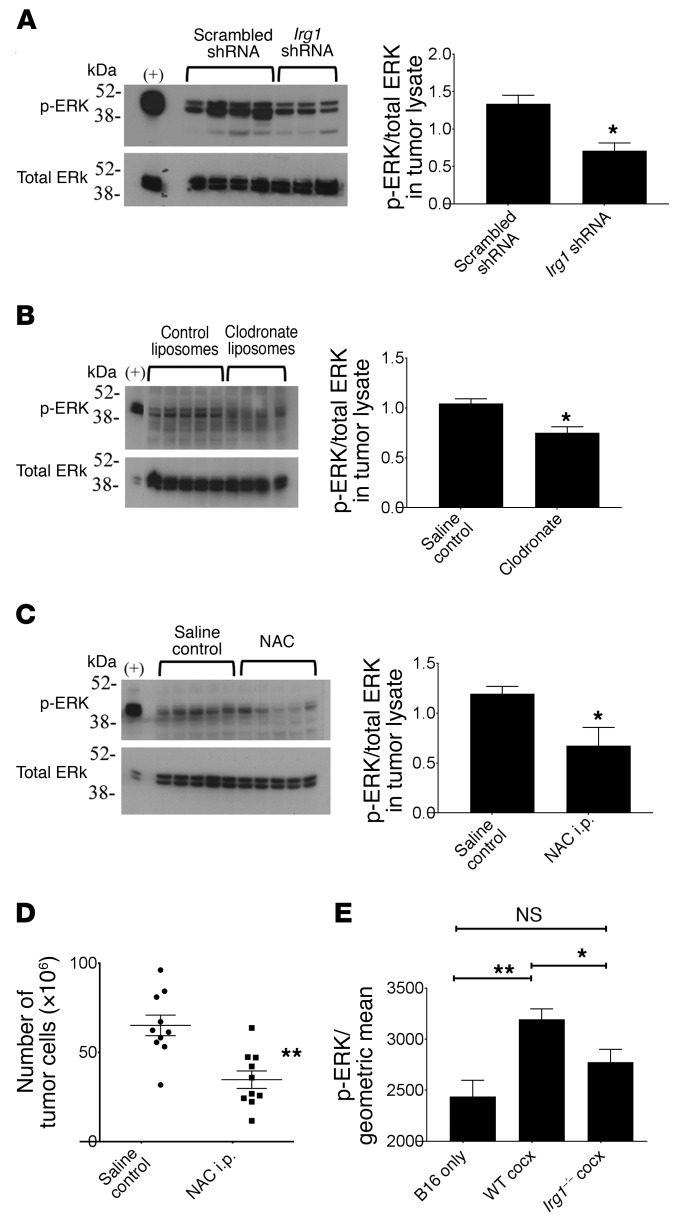
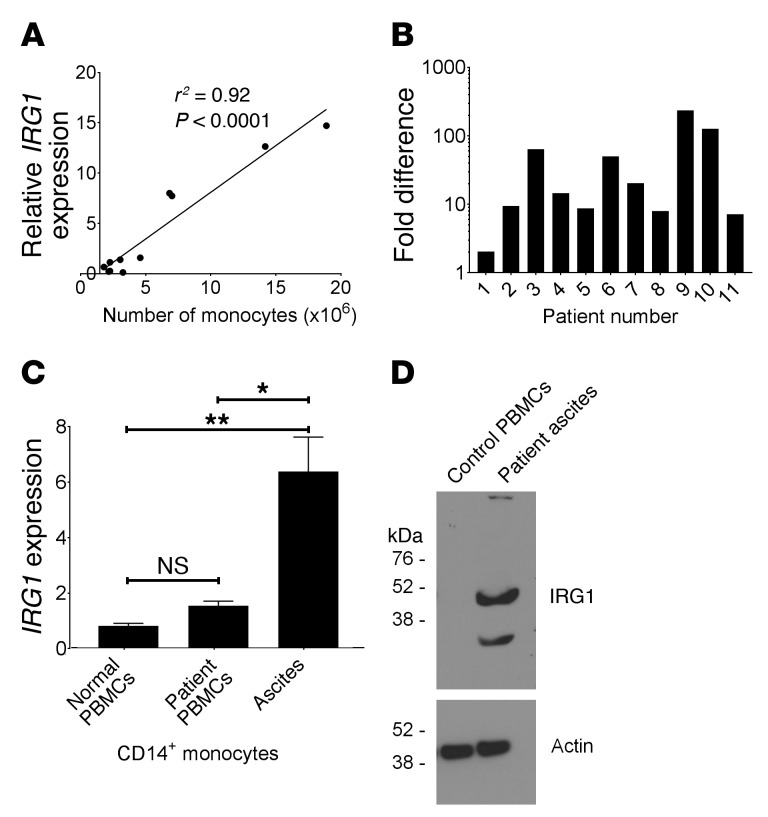
Control of cellular metabolism is critical for efficient cell function, although little is known about the interplay between cell subset-specific metabolites in situ, especially in the tumor setting. Here, we determined how a macrophage-specific (Mϕ-specific) metabolite, itaconic acid, can regulate tumor progression in the peritoneum. We show that peritoneal tumors (B16 melanoma or ID8 ovarian carcinoma) elicited a fatty acid oxidation-mediated increase in oxidative phosphorylation (OXPHOS) and glycolysis in peritoneal tissue-resident macrophages (pResMϕ). Unbiased metabolomics identified itaconic acid, the product of immune-responsive gene 1-mediated (Irg1-mediated) catabolism of mitochondrial cis-aconitate, among the most highly upregulated metabolites in pResMϕ of tumor-bearing mice. Administration of lentivirally encoded Irg1 shRNA significantly reduced peritoneal tumors. This resulted in reductions in OXPHOS and OXPHOS-driven production of ROS in pResMϕ and ROS-mediated MAPK activation in tumor cells. Our findings demonstrate that tumors profoundly alter pResMϕ metabolism, leading to the production of itaconic acid, which potentiates tumor growth. Monocytes isolated from ovarian carcinoma patients' ascites fluid expressed significantly elevated levels of IRG1. Therefore, IRG1 in pResMϕ represents a potential therapeutic target for peritoneal tumors.
Keywords: Intermediary metabolism; Metabolism; Monocytes; Mouse models; Oncology.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
