

Oligomerization of MrgC11 and μ-opioid receptors in sensory neurons enhances morphine analgesia
- PMID: 29921657
- PMCID: PMC6328051
- DOI: 10.1126/scisignal.aao3134
Oligomerization of MrgC11 and μ-opioid receptors in sensory neurons enhances morphine analgesia
Abstract
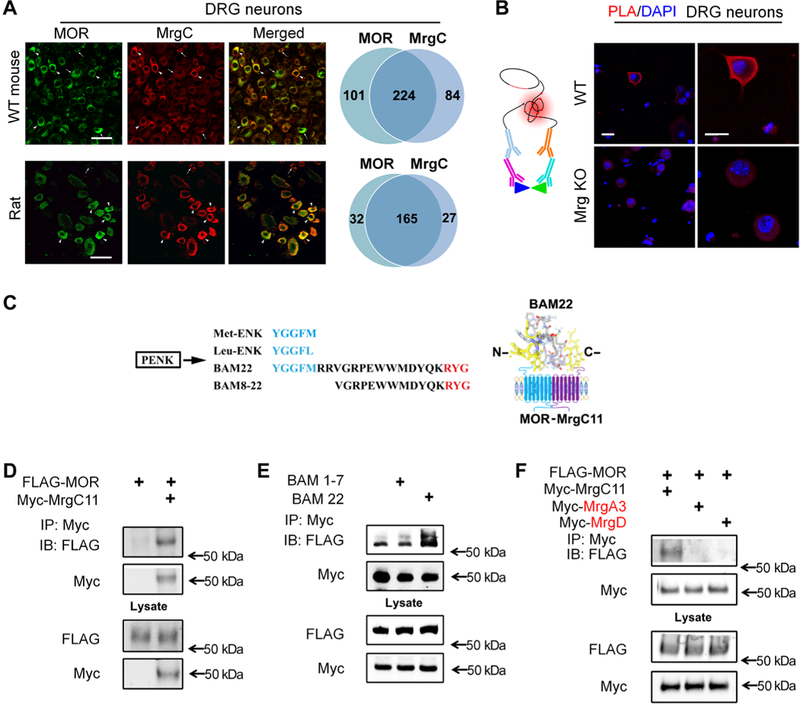
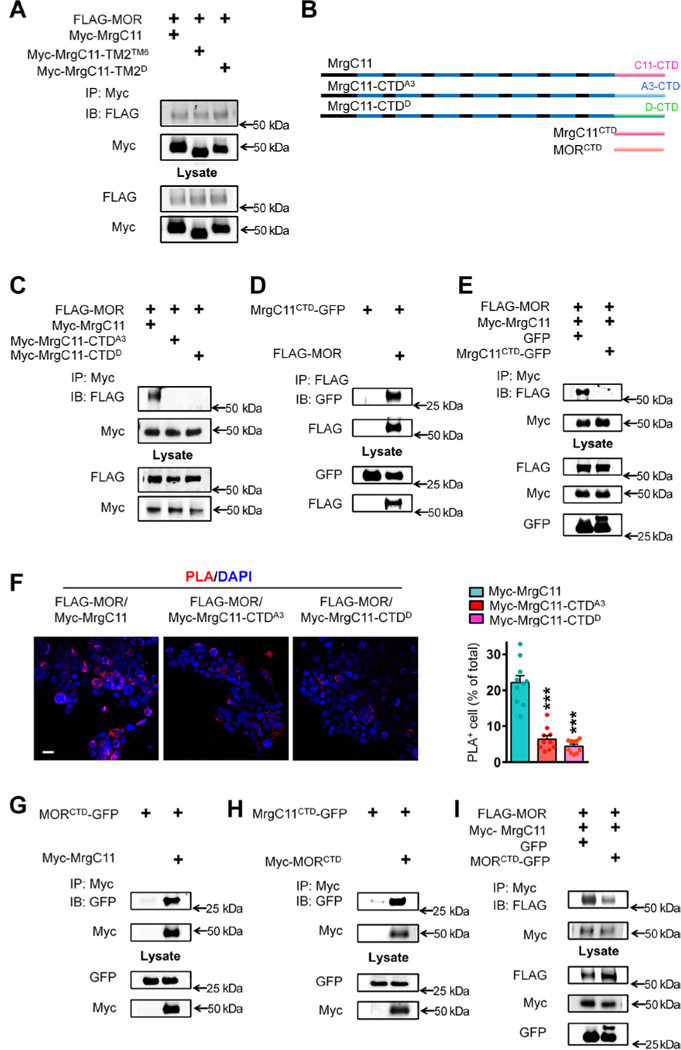
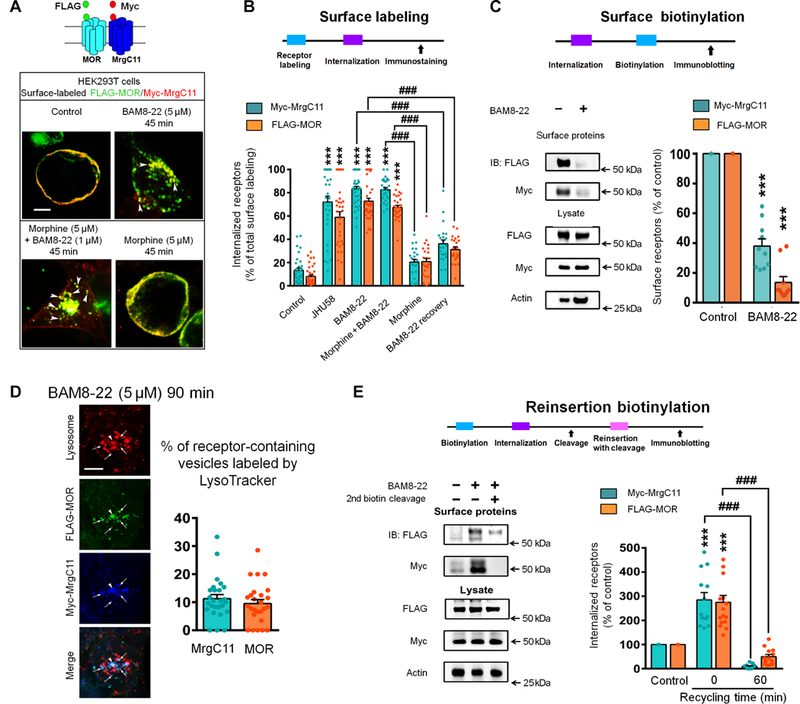
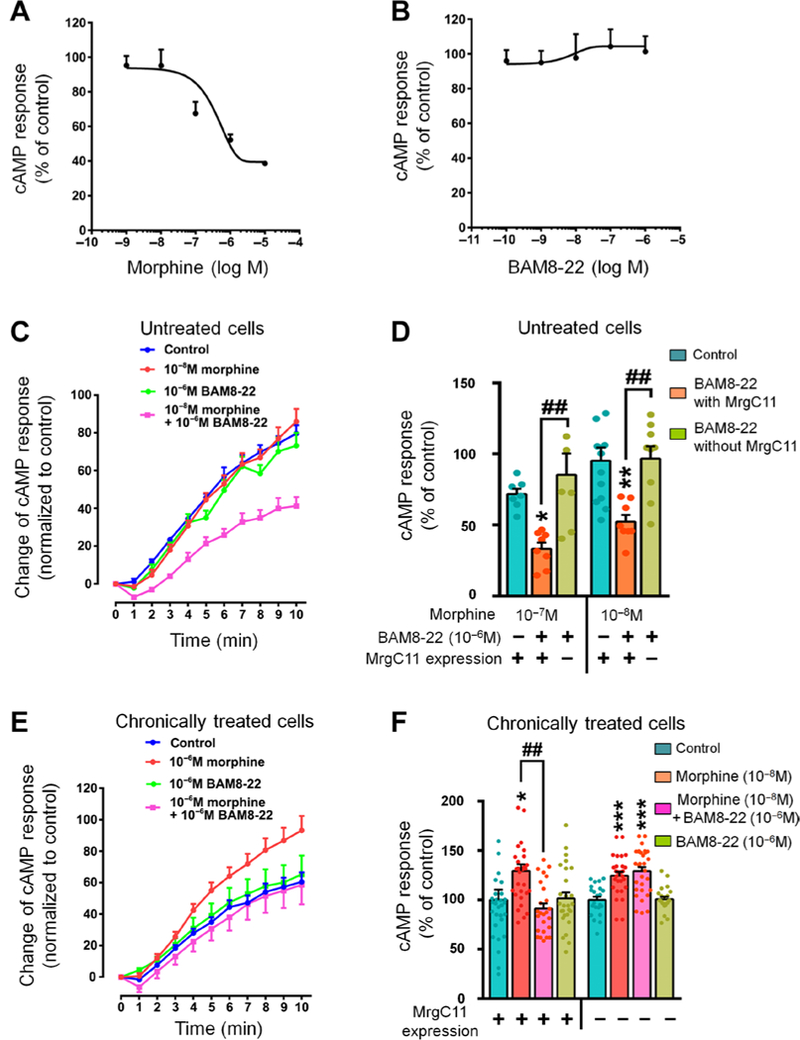
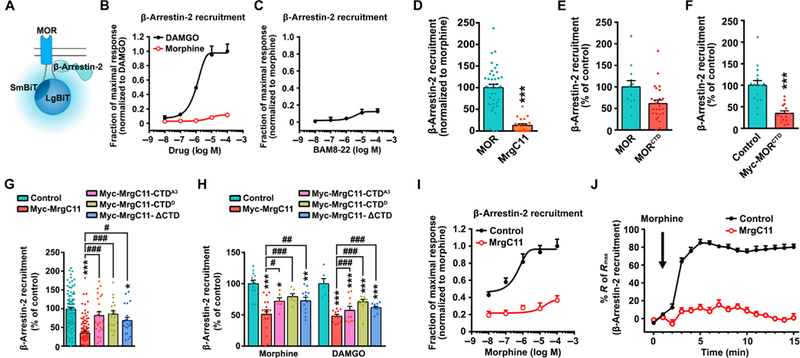
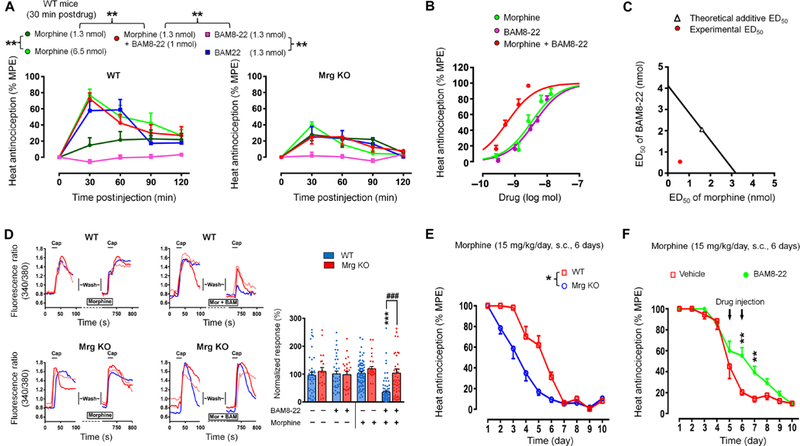
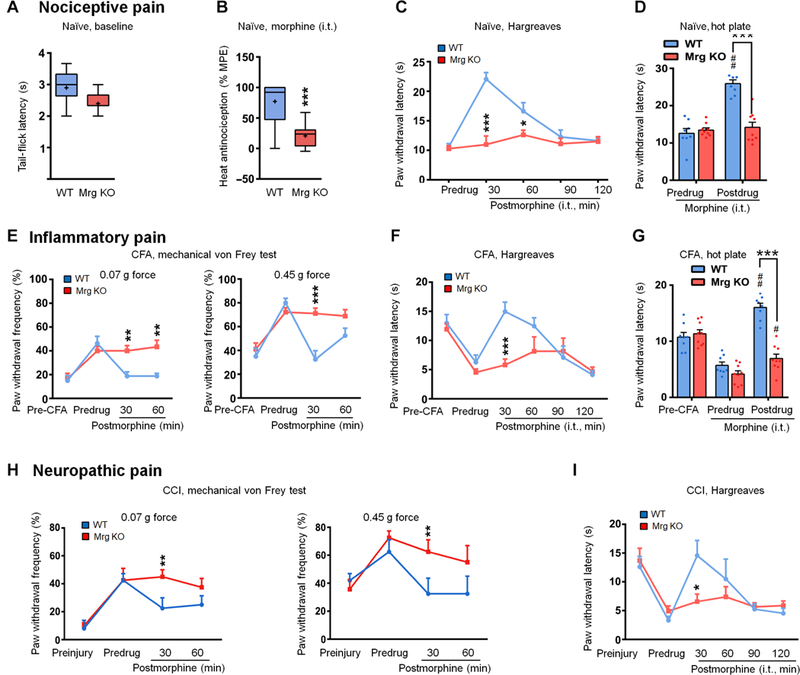
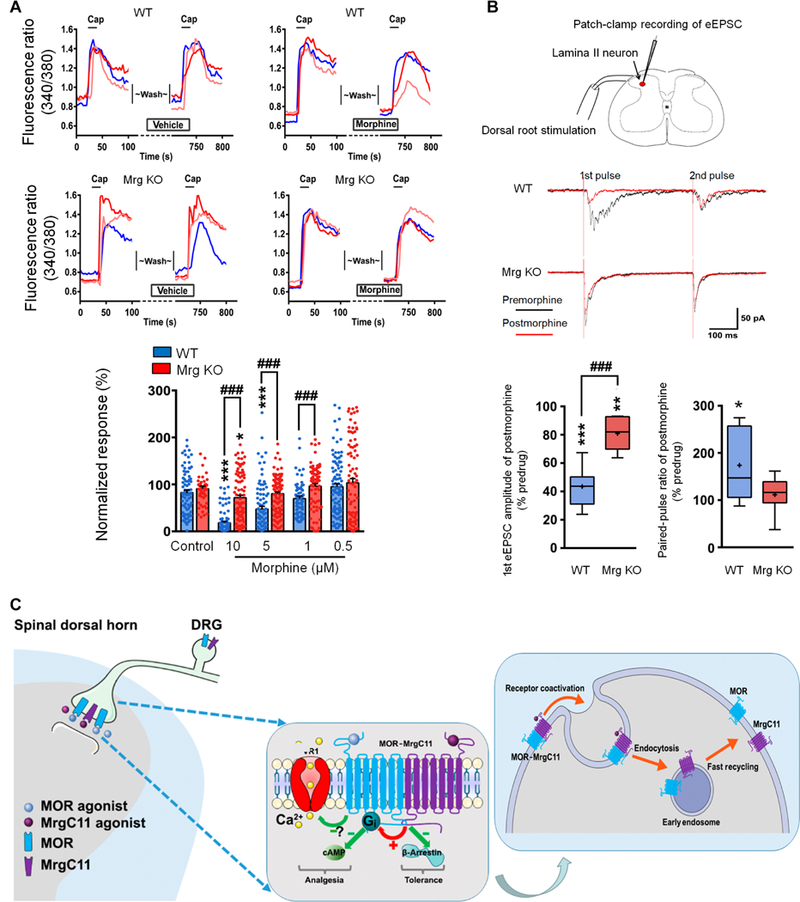
The μ-opioid receptor (MOR) agonist morphine is commonly used for pain management, but it has severe adverse effects and produces analgesic tolerance. Thus, alternative ways of stimulating MOR activity are needed. We found that MrgC11, a sensory neuron-specific G protein-coupled receptor, may form heteromeric complexes with MOR. Peptide-mediated activation of MrgC11 enhanced MOR recycling by inducing coendocytosis and sorting of MOR for membrane reinsertion. MrgC11 activation also inhibited the coupling of MOR to β-arrestin-2 and enhanced the morphine-dependent inhibition of cAMP production. Intrathecal coadministration of a low dose of an MrgC agonist potentiated acute morphine analgesia and reduced chronic morphine tolerance in wild-type mice but not in Mrg-cluster knockout (Mrg KO) mice. BAM22, a bivalent agonist of MrgC and opioid receptors, enhanced the interaction between MrgC11 and MOR and produced stronger analgesia than did the individual monovalent agonists. Morphine-induced neuronal and pain inhibition was reduced in Mrg KO mice compared to that in wild-type mice. Our results uncover MrgC11-MOR interactions that lead to positive functional modulation of MOR. MrgC shares genetic homogeneity and functional similarity with human MrgX1. Thus, harnessing this positive modulation of MOR function by Mrg signaling may enhance morphine analgesia in a sensory neuron-specific fashion to limit central side effects.
Copyright © 2018 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Figures
Similar articles
-
Activation of Mas oncogene-related gene (Mrg) C receptors enhances morphine-induced analgesia through modulation of coupling of μ-opioid receptor to Gi-protein in rat spinal dorsal horn.Neuroscience. 2013 Dec 3;253:455-64. doi: 10.1016/j.neuroscience.2013.08.069. Epub 2013 Sep 14. Neuroscience. 2013. PMID: 24042038
-
Activation of MrgC receptor inhibits N-type calcium channels in small-diameter primary sensory neurons in mice.Pain. 2014 Aug;155(8):1613-1621. doi: 10.1016/j.pain.2014.05.008. Epub 2014 May 9. Pain. 2014. PMID: 24813294 Free PMC article.
-
MrgC agonism at central terminals of primary sensory neurons inhibits neuropathic pain.Pain. 2014 Mar;155(3):534-544. doi: 10.1016/j.pain.2013.12.008. Epub 2013 Dec 11. Pain. 2014. PMID: 24333779 Free PMC article.
-
Ligand-Free Signaling of G-Protein-Coupled Receptors: Relevance to μ Opioid Receptors in Analgesia and Addiction.Molecules. 2022 Sep 8;27(18):5826. doi: 10.3390/molecules27185826. Molecules. 2022. PMID: 36144565 Free PMC article. Review.
-
The mechanism of μ-opioid receptor (MOR)-TRPV1 crosstalk in TRPV1 activation involves morphine anti-nociception, tolerance and dependence.Channels (Austin). 2015;9(5):235-43. doi: 10.1080/19336950.2015.1069450. Epub 2015 Jul 15. Channels (Austin). 2015. PMID: 26176938 Free PMC article. Review.
Cited by
-
The Ubiquitination of Spinal MrgC Alleviates Bone Cancer Pain and Reduces Intracellular Calcium Concentration in Spinal Neurons in Mice.Neurochem Res. 2019 Nov;44(11):2527-2535. doi: 10.1007/s11064-019-02869-3. Epub 2019 Sep 12. Neurochem Res. 2019. PMID: 31515677
-
Role of peripheral sensory neuron mu-opioid receptors in nociceptive, inflammatory, and neuropathic pain.Reg Anesth Pain Med. 2020 Nov;45(11):907-916. doi: 10.1136/rapm-2020-101779. Epub 2020 Sep 14. Reg Anesth Pain Med. 2020. PMID: 32928995 Free PMC article.
-
Investigating G-protein coupled receptor signalling with light-emitting biosensors.Front Physiol. 2024 Jan 8;14:1310197. doi: 10.3389/fphys.2023.1310197. eCollection 2023. Front Physiol. 2024. PMID: 38260094 Free PMC article. Review.
-
Anti-PD-1 treatment impairs opioid antinociception in rodents and nonhuman primates.Sci Transl Med. 2020 Feb 19;12(531):eaaw6471. doi: 10.1126/scitranslmed.aaw6471. Sci Transl Med. 2020. PMID: 32075945 Free PMC article.
-
Discovery of Benzamidine- and 1-Aminoisoquinoline-Based Human MAS-Related G-Protein-Coupled Receptor X1 (MRGPRX1) Agonists.J Med Chem. 2019 Sep 26;62(18):8631-8641. doi: 10.1021/acs.jmedchem.9b01003. Epub 2019 Sep 9. J Med Chem. 2019. PMID: 31498617 Free PMC article.
References
-
- Finnerup NB, Attal N, Haroutounian S, McNicol E, Baron R, Dworkin RH, Gilron I, Haanpaa M, Hansson P, Jensen TS, Kamerman PR, Lund K, Moore A, Raja SN, Rice ASC, Rowbotham M, Sena E, Siddall P, Smith BH, Wallace M, Pharmacotherapy for neuropathic pain in adults: A systematic review and meta-analysis. Lancet Neurol. 14, 162–173 (2015). - PMC - PubMed
-
- He S-Q, Zhang Z-N, Guan J-S, Liu H-R, Zhao B, Wang H-B, Li Q, Yang H, Luo J, Li Z-Y, Wang Q, Lu Y-J, Bao L, Zhang X, Facilitation of μ-opioid receptor activity by preventing 5-opioid receptor-mediated codegradation. Neuron 69, 120–131 (2011). - PubMed
-
- Stein C, Schäfer M, Machelska H, Attacking pain at its source: New perspectives on opioids. Nat. Med. 9, 1003–1008 (2003). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
