

Improving formaldehyde consumption drives methanol assimilation in engineered E. coli
- PMID: 29921903
- PMCID: PMC6008399
- DOI: 10.1038/s41467-018-04795-4
Improving formaldehyde consumption drives methanol assimilation in engineered E. coli
Abstract
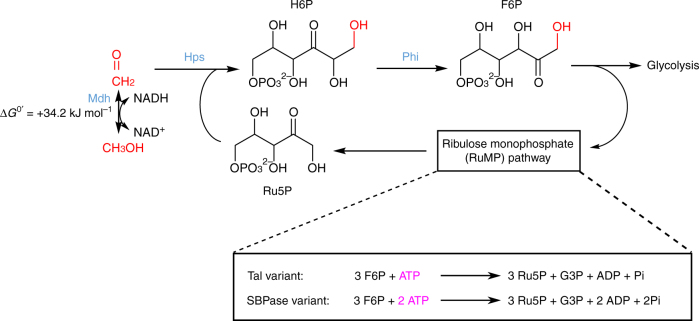
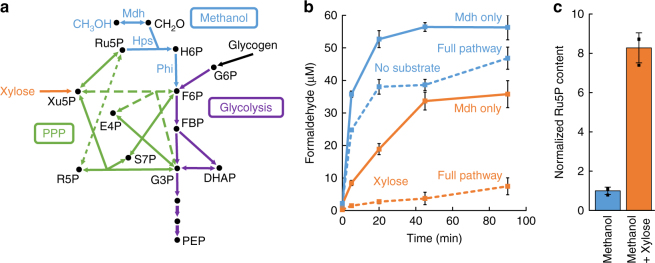
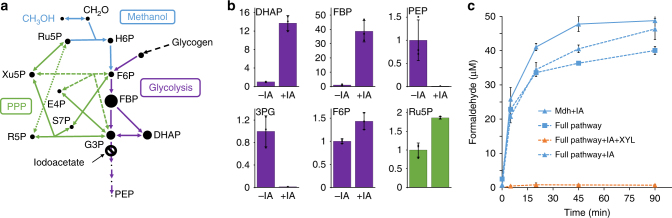
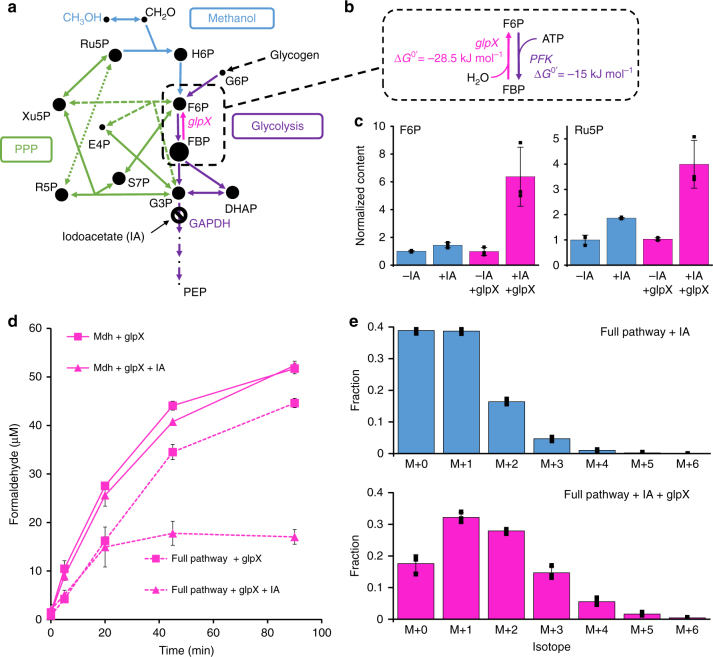
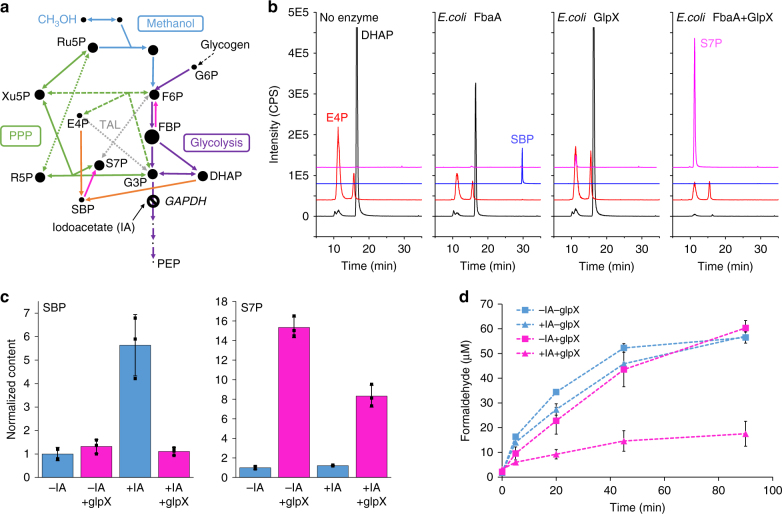
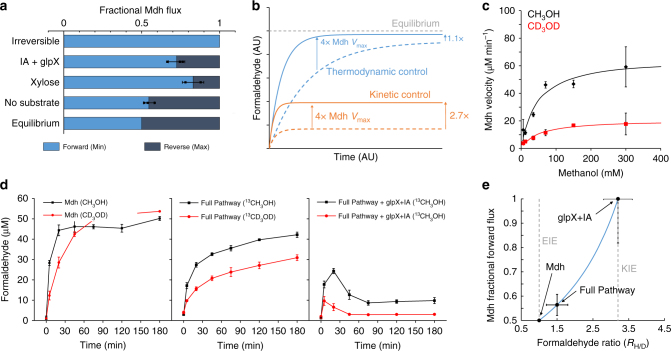
Due to volatile sugar prices, the food vs fuel debate, and recent increases in the supply of natural gas, methanol has emerged as a promising feedstock for the bio-based economy. However, attempts to engineer Escherichia coli to metabolize methanol have achieved limited success. Here, we provide a rigorous systematic analysis of several potential pathway bottlenecks. We show that regeneration of ribulose 5-phosphate in E. coli is insufficient to sustain methanol assimilation, and overcome this by activating the sedoheptulose bisphosphatase variant of the ribulose monophosphate pathway. By leveraging the kinetic isotope effect associated with deuterated methanol as a chemical probe, we further demonstrate that under these conditions overall pathway flux is kinetically limited by methanol dehydrogenase. Finally, we identify NADH as a potent kinetic inhibitor of this enzyme. These results provide direction for future engineering strategies to improve methanol utilization, and underscore the value of chemical biology methodologies in metabolic engineering.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Methanol production by reversed methylotrophy constructed in Escherichia coli.Biosci Biotechnol Biochem. 2020 May;84(5):1062-1068. doi: 10.1080/09168451.2020.1715202. Epub 2020 Jan 16. Biosci Biotechnol Biochem. 2020. PMID: 31942827
-
Engineering the biological conversion of methanol to specialty chemicals in Escherichia coli.Metab Eng. 2017 Jan;39:49-59. doi: 10.1016/j.ymben.2016.10.015. Epub 2016 Nov 1. Metab Eng. 2017. PMID: 27815193
-
Metabolic Engineering of Escherichia coli for High Yield Production of Succinic Acid Driven by Methanol.ACS Synth Biol. 2018 Dec 21;7(12):2803-2811. doi: 10.1021/acssynbio.8b00109. Epub 2018 Nov 28. ACS Synth Biol. 2018. PMID: 30300546
-
Engineering the bioconversion of methane and methanol to fuels and chemicals in native and synthetic methylotrophs.Curr Opin Biotechnol. 2018 Apr;50:81-93. doi: 10.1016/j.copbio.2017.11.010. Epub 2017 Dec 5. Curr Opin Biotechnol. 2018. PMID: 29216497 Review.
-
Recent advances toward the bioconversion of methane and methanol in synthetic methylotrophs.Metab Eng. 2022 May;71:99-116. doi: 10.1016/j.ymben.2021.09.005. Epub 2021 Sep 20. Metab Eng. 2022. PMID: 34547453 Review.
Cited by
-
One-Pot Biosynthesis of 2-Keto-4-hydroxybutyrate from Cheap C1 Compounds Using Rationally Designed Pyruvate Aldolase and Methanol Dehydrogenase.J Agric Food Chem. 2023 Mar 15;71(10):4328-4336. doi: 10.1021/acs.jafc.2c09108. Epub 2023 Mar 1. J Agric Food Chem. 2023. PMID: 36856566 Free PMC article.
-
Sustainable Synthesis of Trimethylolpropane, a Biobased Polyol from Renewable Resources by an Integrated Process of Biotechnology and Chemical Reactions.ACS Omega. 2025 Jul 16;10(29):32413-32423. doi: 10.1021/acsomega.5c04762. eCollection 2025 Jul 29. ACS Omega. 2025. PMID: 40757349 Free PMC article.
-
Improving the Methanol Tolerance of an Escherichia coli Methylotroph via Adaptive Laboratory Evolution Enhances Synthetic Methanol Utilization.Front Microbiol. 2021 Feb 11;12:638426. doi: 10.3389/fmicb.2021.638426. eCollection 2021. Front Microbiol. 2021. PMID: 33643274 Free PMC article.
-
Methanol-dependent Escherichia coli strains with a complete ribulose monophosphate cycle.Nat Commun. 2020 Oct 26;11(1):5403. doi: 10.1038/s41467-020-19235-5. Nat Commun. 2020. PMID: 33106470 Free PMC article.
-
Evolutionary engineering of methylotrophic E. coli enables fast growth on methanol.Nat Commun. 2024 Oct 13;15(1):8840. doi: 10.1038/s41467-024-53206-4. Nat Commun. 2024. PMID: 39397031 Free PMC article.
References
-
- Clomburg, J. M., Crumbley, A. M. & Gonzalez, R. Industrial biomanufacturing: the future of chemical production. Science355, aag0804 (2017). - PubMed
-
- Paré, A. Enerkem’s facility in Edmonton becomes the first ISCC certified plant in the world to convert municipal solid waste into biomethanol. Enerkem http://enerkem.com/newsroom/?communique_id=122557 (2016)
-
- CRI. National Planning Agency approves environmental impact assessment for CRI’s CO2-to-methanol plant. Carbon Recycling International http://carbonrecycling.is/news/2017/5/22/national-planning-agency-approv... (2017).
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
