

PDCD5 regulates iNKT cell terminal maturation and iNKT1 fate decision
- PMID: 29921968
- PMCID: PMC6804728
- DOI: 10.1038/s41423-018-0059-2
PDCD5 regulates iNKT cell terminal maturation and iNKT1 fate decision
Abstract
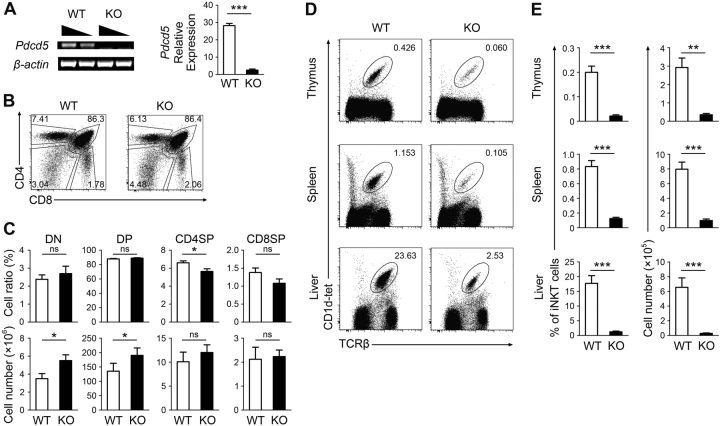
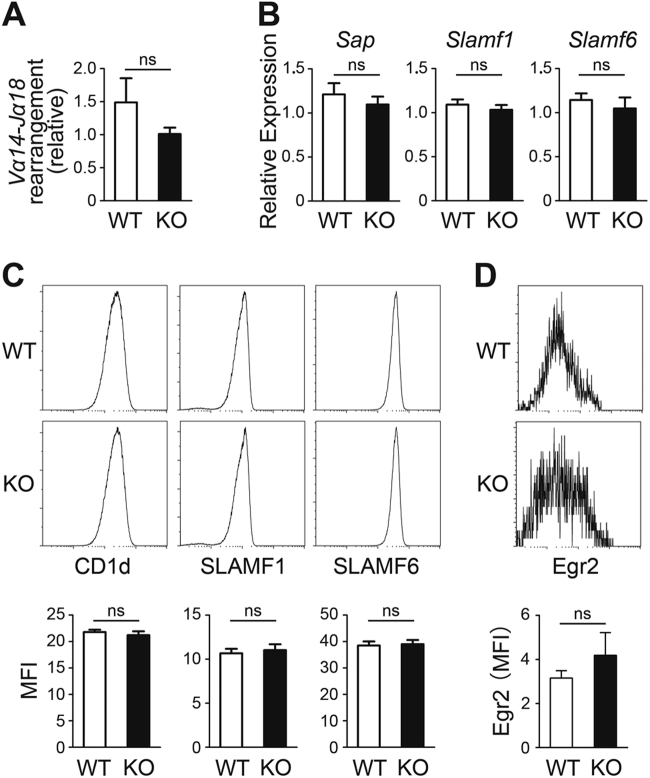
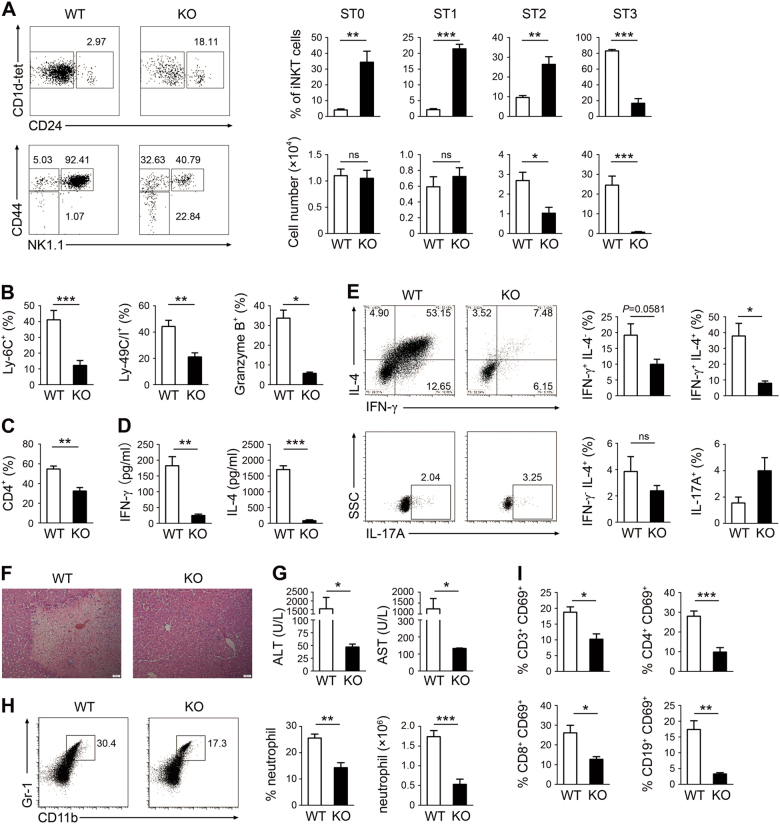
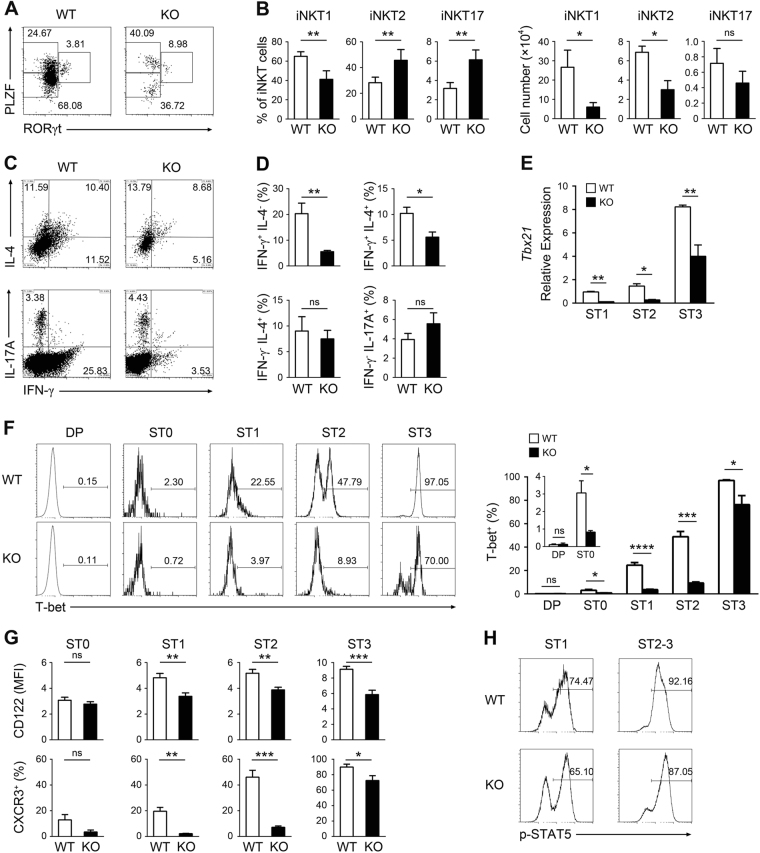
Invariant natural killer T1 (iNKT1) cells are characterized by the preferential expression of T-box transcription factor T-bet (encoded by Tbx21) and the production of cytokine IFN-γ, but the relationship between the developmental process and iNKT1 lineage diversification in the thymus remains elusive. We report in the present study a crucial role of programmed cell death 5 (PDCD5) in iNKT cell terminal maturation and iNKT1 fate determination. Mice with T cell-specific deletion of PDCD5 had decreased numbers of thymic and peripheral iNKT cells with a predominantly immature phenotype and defects in response to α-galactosylceramide. Loss of PDCD5 also selectively abolished the iNKT1 lineage by reducing T-bet expression in iNKT cells at an early thymic developmental stage (before CD44 upregulation). We further demonstrated that TOX2, one of the high mobility group proteins that was highly expressed in iNKT cells at stage 1 and could be stabilized by PDCD5, promoted the permissive histone H3K4me3 modification in the promoter region of Tbx21. These data indicate a pivotal and unique role of PDCD5/TOX2 in iNKT1 lineage determination. They also suggest that the fate of iNKT1 may be programmed at the developmental stage of iNKT cells in the thymus.
Conflict of interest statement
The authors declare no competing interests.
Figures



References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
