

COI1-dependent jasmonate signalling affects growth, metabolite production and cell wall protein composition in arabidopsis
- PMID: 29924303
- PMCID: PMC6324744
- DOI: 10.1093/aob/mcy109
COI1-dependent jasmonate signalling affects growth, metabolite production and cell wall protein composition in arabidopsis
Abstract
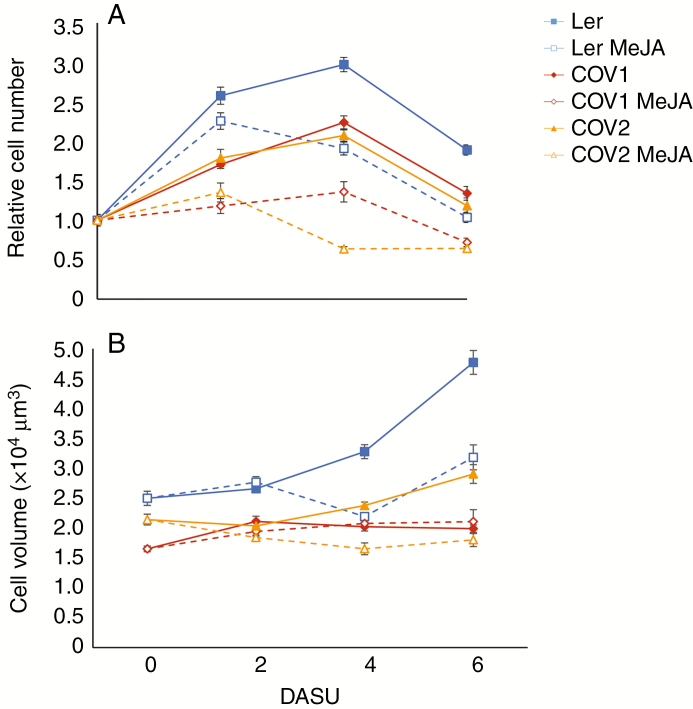
Background and aims: Cultured cell suspensions have been the preferred model to study the apoplast as well as to monitor metabolic and cell cycle-related changes. Previous work showed that methyl jasmonate (MeJA) inhibits leaf growth in a CORONATINE INSENSITIVE 1 (COI1)-dependent manner, with COI1 being the jasmonate (JA) receptor. Here, the effect of COI1 overexpression on the growth of stably transformed arabidopsis cell cultures is described.
Methods: Time-course experiments were carried out to analyse gene expression, and protein and metabolite levels.
Key results: Both MeJA treatment and the overexpression of COI1 modify growth, by altering cell proliferation and expansion. DNA content as well as transcript patterns of cell cycle and cell wall remodelling markers were altered. COI1 overexpression also increases the protein levels of OLIGOGALACTURONIDE OXIDASE 1, BETA-GLUCOSIDASE/ENDOGLUCANASES and POLYGALACTURONASE INHIBITING PROTEIN2, reinforcing the role of COI1 in mediating defence responses and highlighting a link between cell wall loosening and growth regulation. Moreover, changes in the levels of the primary metabolites alanine, serine and succinic acid of MeJA-treated Arabidopsis cell cultures were observed. In addition, COI1 overexpression positively affects the availability of metabolites such as β-alanine, threonic acid, putrescine, glucose and myo-inositol, thereby providing a connection between JA-inhibited growth and stress responses.
Conclusions: This study contributes to the understanding of the regulation of growth and the production of metabolic resources by JAs and COI1. This will have important implications in dissecting the complex relationships between hormonal and cell wall signalling in plants. The work also provides tools to uncover novel mechanisms co-ordinating cell division and post-mitotic cell expansion in the absence of organ developmental control.
Figures






References
-
- Akiyama T, Jin S, Yoshida M, Hoshino T, Opassiri R, Cairns JRK. 2009. Expression of an endo-(1,3;1,4)-β-glucanase in response to wounding, methyl jasmonate, abscisic acid and ethephon in rice seedlings. Journal of Plant Physiology 166: 1814–1825. - PubMed
-
- An SH, Choi HW, Hong JK, Hwang BK. 2009. Regulation and function of the pepper pectin methylesterase inhibitor (CaPMEI1) gene promoter in defense and ethylene and methyl jasmonate signaling in plants. Planta 230: 1223–1237. - PubMed
-
- Axelos M, Curie C, Mazzolini L, Bardet C, Lescure B. 1992. A protocol for transient gene expression in Arabidopsis thaliana protoplasts isolated from cell-suspension cultures. Plant Physiology and Biochemistry 30: 123–128.
-
- Balbi V, Devoto A. 2008. Jasmonate signalling network in Arabidopsis thaliana: crucial regulatory nodes and new physiological scenarios. New Phytologist 177: 301–318. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
