

In silico and ex vivo approaches indicate immune pressure on capsid and non-capsid regions of coxsackie B viruses in the human system
- PMID: 29924875
- PMCID: PMC6010236
- DOI: 10.1371/journal.pone.0199323
In silico and ex vivo approaches indicate immune pressure on capsid and non-capsid regions of coxsackie B viruses in the human system
Abstract
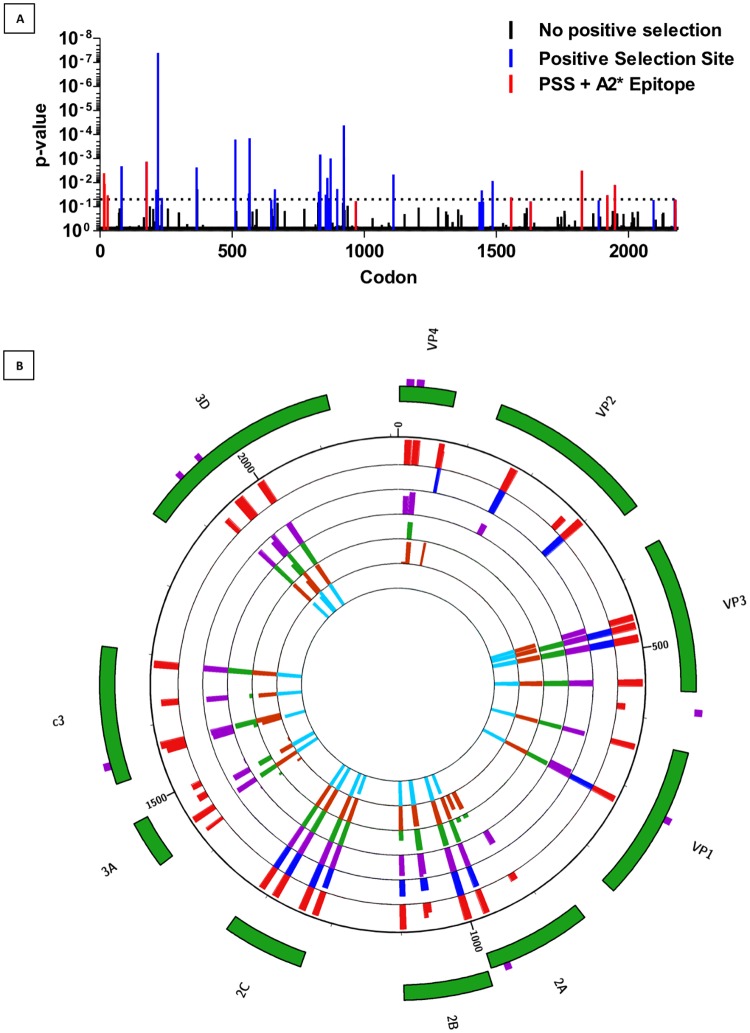
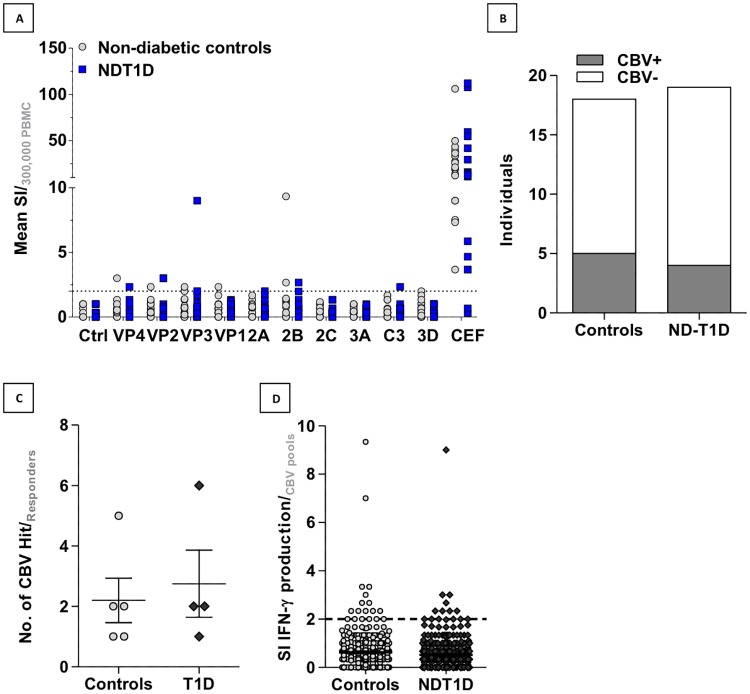
Coxsackie B Virus (CBV) infection has been linked to the aetiology of type 1 diabetes (T1D) and vaccination has been proposed as prophylaxis for disease prevention. Serum neutralising antibodies and the presence of viral protein and RNA in tissues have been common tools to examine this potential disease relationship, whilst the role of anti-CBV cytotoxic T cell responses and their targets have not been studied. To address this knowledge gap, we augmented conventional HLA-binding predictive algorithm-based epitope discovery by cross-referencing epitopes with sites of positive natural selection within the CBV3 viral genome, identified using mixed effects models of evolution. Eight epitopes for the common MHC class I allele HLA-A*0201 occur at sites that appear to be positively selected. Furthermore, such epitopes span the viral genome, indicating that effective anti-viral responses may not be restricted to the capsid region. To assess the spectrum of IFNy responses in non-diabetic subjects and recently diagnosed type 1 diabetes (T1D) patients, we stimulated PBMC ex vivo with pools of synthetic peptides based on component-restricted sequences identified in silico. We found responders were more likely to recognize multiple rather than a single CBV peptide pool, indicating that the natural course of infection results in multiple targets for effector memory responses, rather than immunodominant epitopes or viral components. The finding that anti-CBV CD8 T cell immunity is broadly targeted has implications for vaccination strategies and studies on the pathogenesis of CBV-linked diseases.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
A study of the cellular immune response to enteroviruses in humans: identification of cross-reactive T cell epitopes on the structural proteins of enteroviruses.J Gen Virol. 1996 Sep;77 ( Pt 9):2097-108. doi: 10.1099/0022-1317-77-9-2097. J Gen Virol. 1996. PMID: 8811009
-
Identification of serotype-specific and nonserotype-specific B-cell epitopes of coxsackie B virus using synthetic peptides.Virology. 1994 May 1;200(2):381-9. doi: 10.1006/viro.1994.1202. Virology. 1994. PMID: 7513917
-
In silico analysis of MHC-I restricted epitopes of Chikungunya virus proteins: Implication in understanding anti-CHIKV CD8(+) T cell response and advancement of epitope based immunotherapy for CHIKV infection.Infect Genet Evol. 2015 Apr;31:118-26. doi: 10.1016/j.meegid.2015.01.017. Epub 2015 Jan 31. Infect Genet Evol. 2015. PMID: 25643869
-
Amino Acid Substitutions within HLA-B*27-Restricted T Cell Epitopes Prevent Recognition by Hepatitis Delta Virus-Specific CD8+ T Cells.J Virol. 2018 Jun 13;92(13):e01891-17. doi: 10.1128/JVI.01891-17. Print 2018 Jul 1. J Virol. 2018. PMID: 29669837 Free PMC article.
-
[MHC tetramers: tracking specific immunity].Acta Med Croatica. 2003;57(4):255-9. Acta Med Croatica. 2003. PMID: 14639858 Review. Croatian.
Cited by
-
Pancreas Pathology During the Natural History of Type 1 Diabetes.Curr Diab Rep. 2018 Oct 6;18(11):124. doi: 10.1007/s11892-018-1084-3. Curr Diab Rep. 2018. PMID: 30293191 Review.
-
Rationale for enteroviral vaccination and antiviral therapies in human type 1 diabetes.Diabetologia. 2019 May;62(5):744-753. doi: 10.1007/s00125-019-4811-7. Epub 2019 Jan 23. Diabetologia. 2019. PMID: 30675626 Free PMC article. Review.
-
Enterovirus infection and type 1 diabetes: unraveling the crime scene.Clin Exp Immunol. 2019 Jan;195(1):15-24. doi: 10.1111/cei.13223. Epub 2018 Nov 13. Clin Exp Immunol. 2019. PMID: 30307605 Free PMC article. Review.
-
Genetic and Environmental Interaction in Type 1 Diabetes: a Relationship Between Genetic Risk Alleles and Molecular Traits of Enterovirus Infection?Curr Diab Rep. 2019 Aug 10;19(9):82. doi: 10.1007/s11892-019-1192-8. Curr Diab Rep. 2019. PMID: 31401790 Free PMC article. Review.
References
-
- Oikarinen S, Tauriainen S, Hober D, Lucas B, Vazeou A, Sioofy-Khojine A, et al. Virus antibody survey in different european populations indicates risk association between coxsackievirus B1 and type 1 diabetes. Diabetes. 2014;63: 655–662. doi: 10.2337/db13-0620 - DOI - PubMed
-
- Chehadeh W, Weill J, Vantyghem MC, Alm G, Lefèbvre J, Wattré P, et al. Increased level of interferon-alpha in blood of patients with insulin-dependent diabetes mellitus: relationship with coxsackievirus B infection. J Infect Dis. 2000;181: 1929–1939. doi: 10.1086/315516 - DOI - PubMed
-
- Lin H-C, Wang C-H, Tsai F-J, Hwang K-P, Chen W, Lin C-C, et al. Enterovirus infection is associated with an increased risk of childhood type 1 diabetes in Taiwan: a nationwide population-based cohort study. Diabetologia. 2015;58: 79–86. doi: 10.1007/s00125-014-3400-z - DOI - PubMed
-
- Richardson SJ, Willcox a., Bone a. J, Foulis a. K, Morgan NG. The prevalence of enteroviral capsid protein vp1 immunostaining in pancreatic islets in human type 1 diabetes. Diabetologia. 2009;52: 1143–1151. doi: 10.1007/s00125-009-1276-0 - DOI - PubMed
-
- Yin H, Berg A-K, Tuvemo T, Frisk G. Enterovirus RNA is found in peripheral blood mononuclear cells in a majority of type 1 diabetic children at onset. Diabetes. 2002;51: 1964–1971. doi: 10.2337/diabetes.51.6.1964 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
