

How Messenger RNA and Nascent Chain Sequences Regulate Translation Elongation
- PMID: 29925264
- PMCID: PMC6594189
- DOI: 10.1146/annurev-biochem-060815-014818
How Messenger RNA and Nascent Chain Sequences Regulate Translation Elongation
Abstract
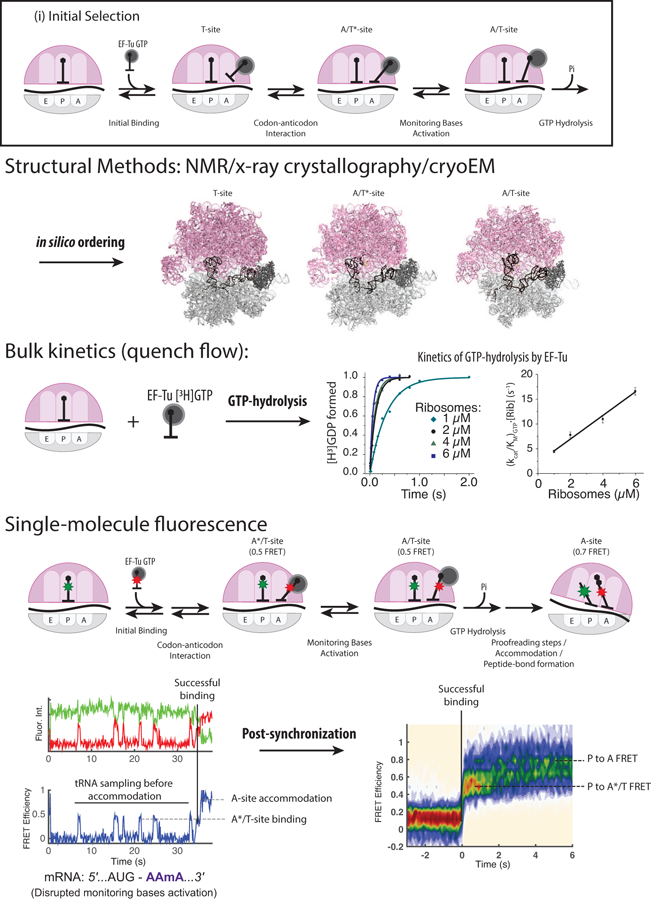
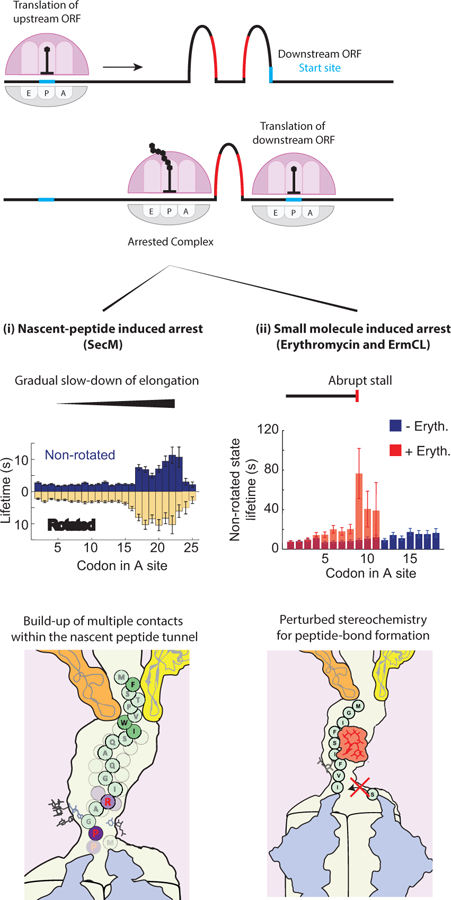
Translation elongation is a highly coordinated, multistep, multifactor process that ensures accurate and efficient addition of amino acids to a growing nascent-peptide chain encoded in the sequence of translated messenger RNA (mRNA). Although translation elongation is heavily regulated by external factors, there is clear evidence that mRNA and nascent-peptide sequences control elongation dynamics, determining both the sequence and structure of synthesized proteins. Advances in methods have driven experiments that revealed the basic mechanisms of elongation as well as the mechanisms of regulation by mRNA and nascent-peptide sequences. In this review, we highlight how mRNA and nascent-peptide elements manipulate the translation machinery to alter the dynamics and pathway of elongation.
Keywords: mRNA; nascent-peptide chain; protein synthesis; recoding; ribosome; translation control.
Figures



References
-
- Atkins JF, Gesteland RF, eds. 2010. Recoding: Expansion of Decoding Rules Enriches Gene Expression New: Springer Verlag
-
- Ito K, Chiba S. 2013. Arrest Peptides: Cis -Acting Modulators of Translation. Annu. Rev. Biochem 82(1):171–202 - PubMed
-
- Wilson DN, Arenz S, Beckmann R. 2016. Translation regulation via nascent polypeptide-mediated ribosome stalling. Curr. Opin. Struct. Biol 37:123–33 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
