

Secretory Vesicle Polar Sorting, Endosome Recycling and Cytoskeleton Organization Require the AP-1 Complex in Aspergillus nidulans
- PMID: 29925567
- PMCID: PMC6063236
- DOI: 10.1534/genetics.118.301240
Secretory Vesicle Polar Sorting, Endosome Recycling and Cytoskeleton Organization Require the AP-1 Complex in Aspergillus nidulans
Abstract
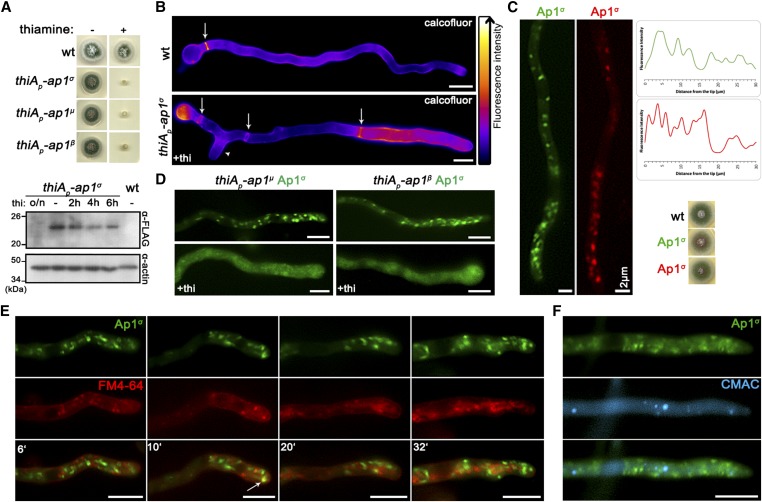
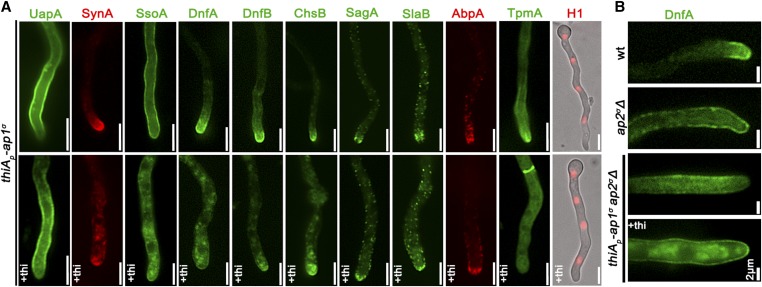
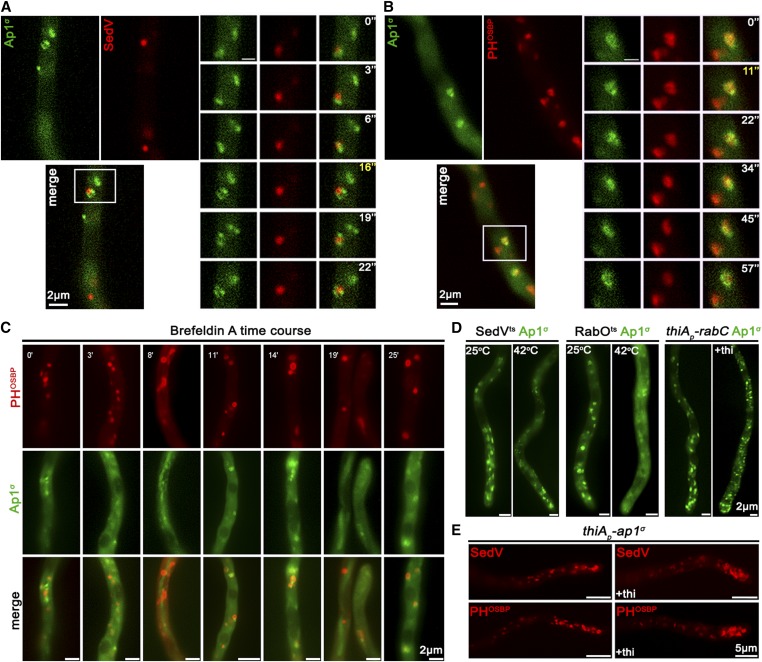
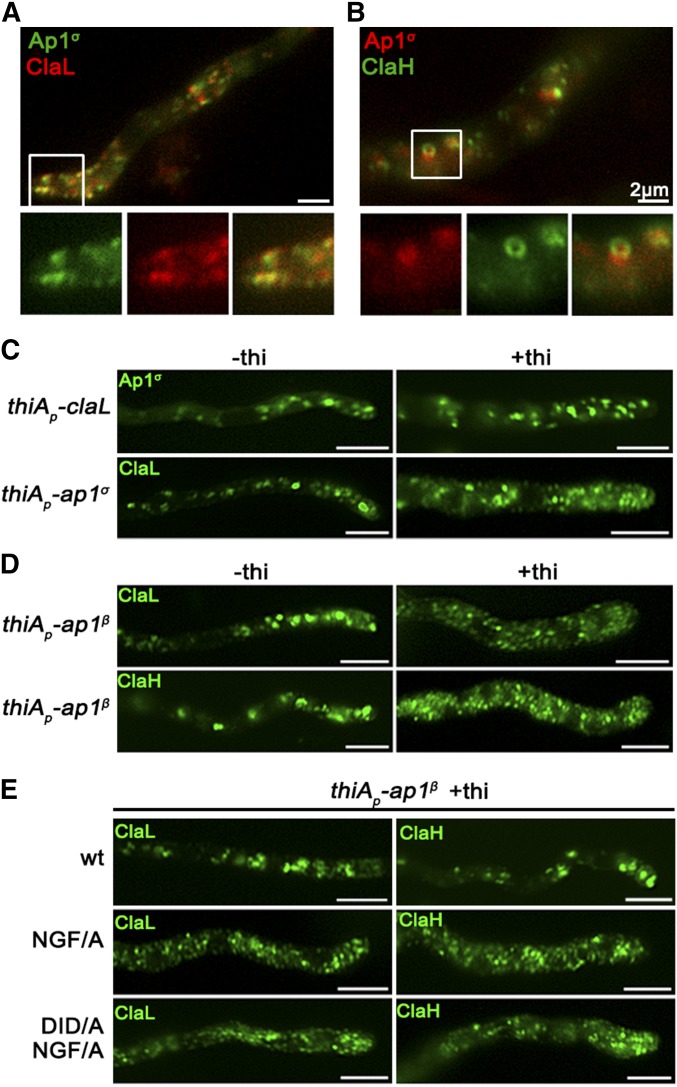
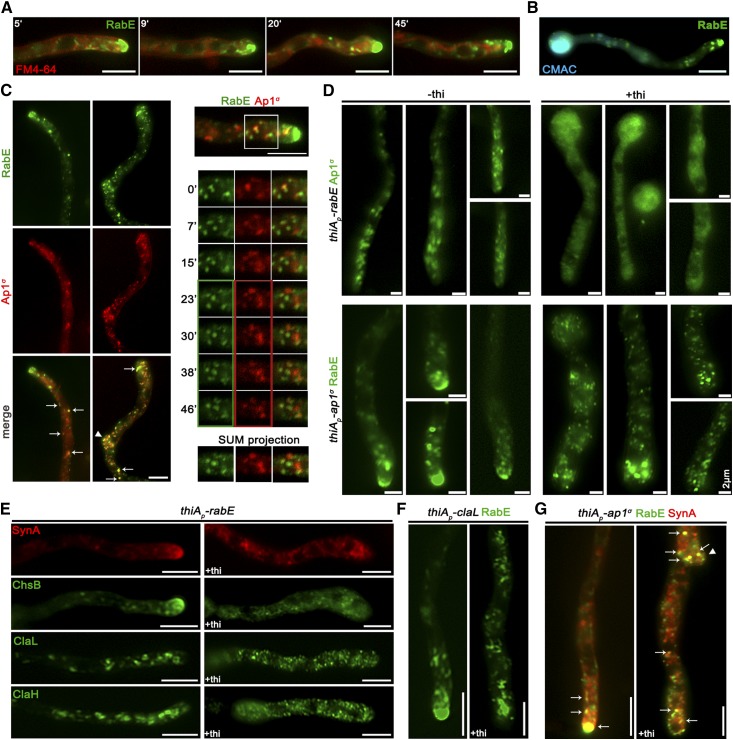
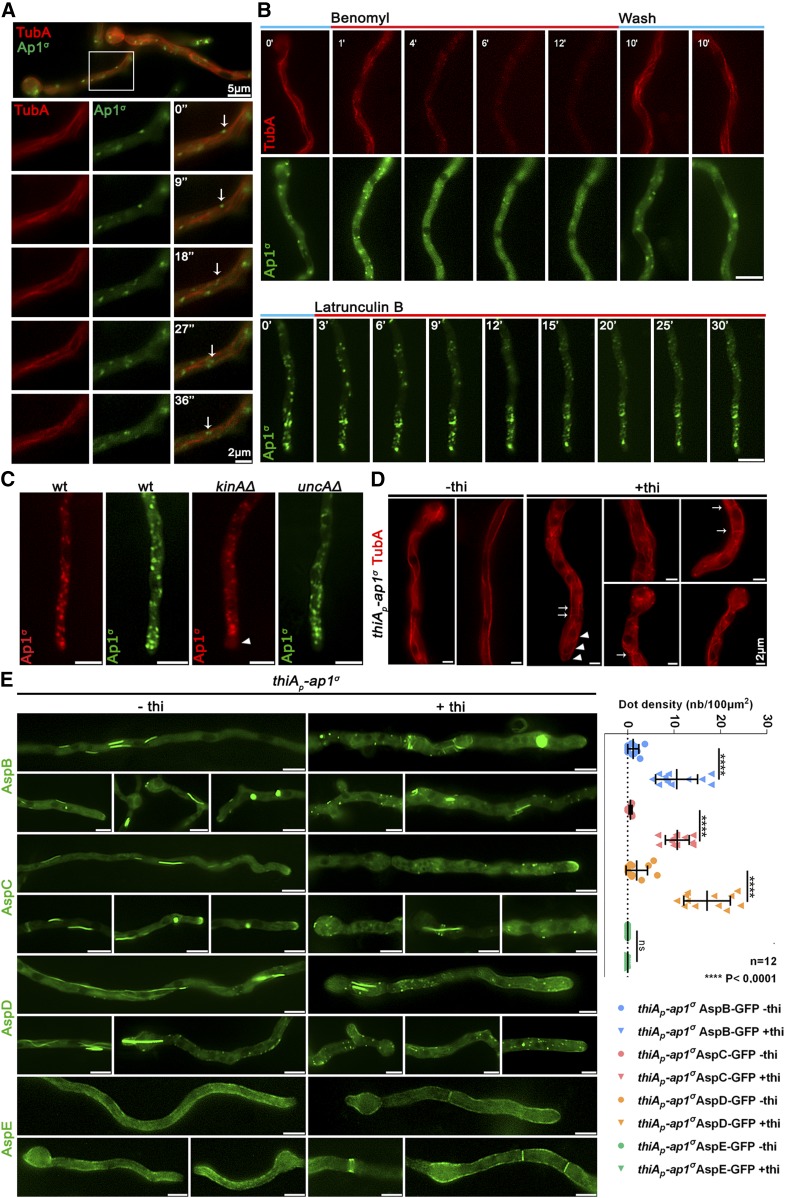
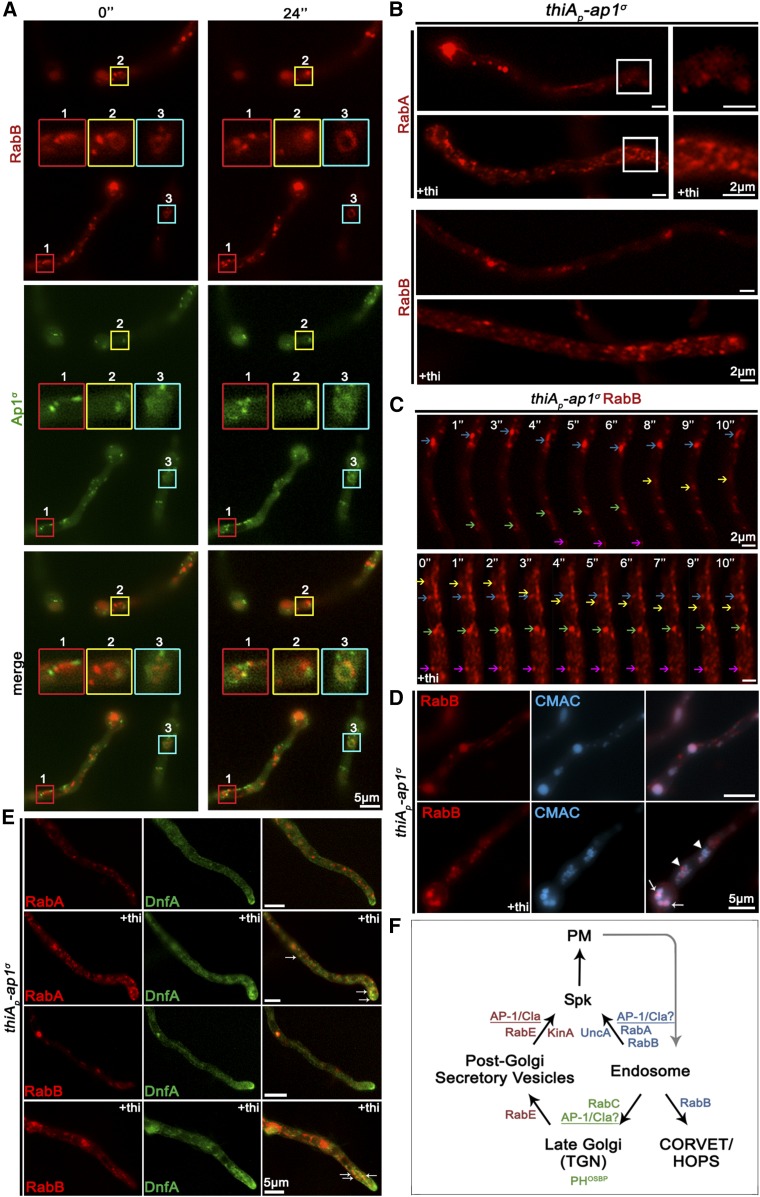
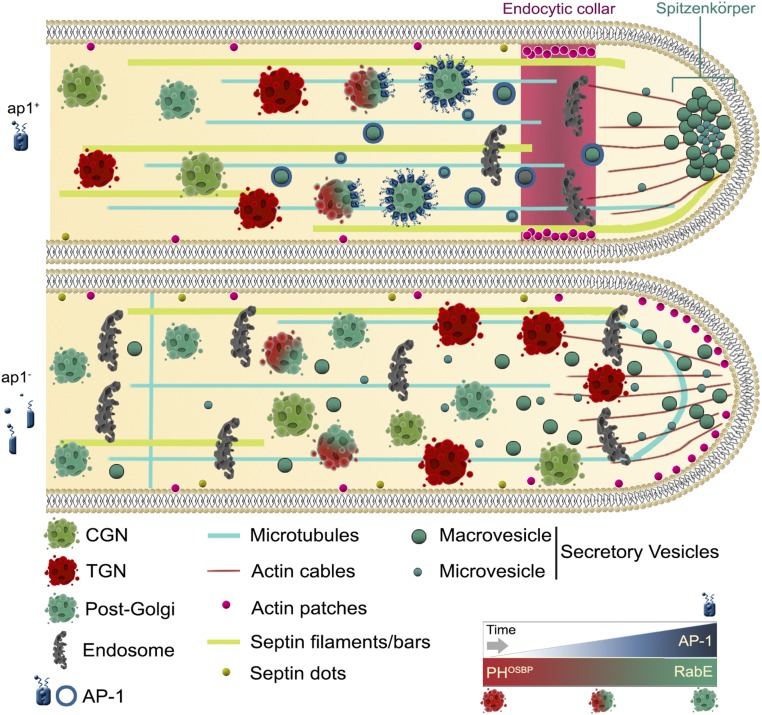
The AP-1 complex is essential for membrane protein traffic via its role in the pinching-off and sorting of secretory vesicles (SVs) from the trans-Golgi and/or endosomes. While its essentiality is undisputed in metazoa, its role in simpler eukaryotes seems less clear. Here, we dissect the role of AP-1 in the filamentous fungus Aspergillus nidulans and show that it is absolutely essential for growth due to its role in clathrin-dependent maintenance of polar traffic of specific membrane cargoes toward the apex of growing hyphae. We provide evidence that AP-1 is involved in both anterograde sorting of RabERab11-labeled SVs and RabA/BRab5-dependent endosome recycling. Additionally, AP-1 is shown to be critical for microtubule and septin organization, further rationalizing its essentiality in cells that face the challenge of cytoskeleton-dependent polarized cargo traffic. This work also opens a novel issue on how nonpolar cargoes, such as transporters, are sorted to the eukaryotic plasma membrane.
Keywords: Rab GTPases; fungi; microtubules; secretion; traffic; transport.
Copyright © 2018 by the Genetics Society of America.
Figures
Similar articles
-
Translocation of nutrient transporters to cell membrane via Golgi bypass in Aspergillus nidulans.EMBO Rep. 2020 Jul 3;21(7):e49929. doi: 10.15252/embr.201949929. Epub 2020 May 26. EMBO Rep. 2020. PMID: 32452614 Free PMC article.
-
Transport of fungal RAB11 secretory vesicles involves myosin-5, dynein/dynactin/p25, and kinesin-1 and is independent of kinesin-3.Mol Biol Cell. 2017 Apr 1;28(7):947-961. doi: 10.1091/mbc.E16-08-0566. Epub 2017 Feb 16. Mol Biol Cell. 2017. PMID: 28209731 Free PMC article.
-
Life and Death of Fungal Transporters under the Challenge of Polarity.Int J Mol Sci. 2020 Jul 29;21(15):5376. doi: 10.3390/ijms21155376. Int J Mol Sci. 2020. PMID: 32751072 Free PMC article. Review.
-
The Golgi apparatus: insights from filamentous fungi.Mycologia. 2016 May-Jun;108(3):603-22. doi: 10.3852/15-309. Epub 2016 Mar 1. Mycologia. 2016. PMID: 26932185 Review.
-
The AP-2 complex has a specialized clathrin-independent role in apical endocytosis and polar growth in fungi.Elife. 2017 Feb 21;6:e20083. doi: 10.7554/eLife.20083. Elife. 2017. PMID: 28220754 Free PMC article.
Cited by
-
ADAPTOR PROTEIN-1 complex-mediated post-Golgi trafficking is critical for pollen wall development in Arabidopsis.New Phytol. 2022 Jul;235(2):472-487. doi: 10.1111/nph.18170. Epub 2022 May 21. New Phytol. 2022. PMID: 35451504 Free PMC article.
-
Translocation of nutrient transporters to cell membrane via Golgi bypass in Aspergillus nidulans.EMBO Rep. 2020 Jul 3;21(7):e49929. doi: 10.15252/embr.201949929. Epub 2020 May 26. EMBO Rep. 2020. PMID: 32452614 Free PMC article.
-
Distinct trafficking routes of polarized and non-polarized membrane cargoes in Aspergillus nidulans.Elife. 2024 Oct 21;13:e103355. doi: 10.7554/eLife.103355. Elife. 2024. PMID: 39431919 Free PMC article.
-
New directions for the clathrin adaptor AP-1 in cell biology and human disease.Curr Opin Cell Biol. 2022 Jun;76:102079. doi: 10.1016/j.ceb.2022.102079. Epub 2022 Apr 13. Curr Opin Cell Biol. 2022. PMID: 35429729 Free PMC article. Review.
-
Transmembrane helices 5 and 12 control transport dynamics, substrate affinity, and specificity in the elevator-type UapA transporter.Genetics. 2022 Aug 30;222(1):iyac107. doi: 10.1093/genetics/iyac107. Genetics. 2022. PMID: 35894659 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
