

Consequences of cathepsin C inactivation for membrane exposure of proteinase 3, the target antigen in autoimmune vasculitis
- PMID: 29925593
- PMCID: PMC6093229
- DOI: 10.1074/jbc.RA118.001922
Consequences of cathepsin C inactivation for membrane exposure of proteinase 3, the target antigen in autoimmune vasculitis
Abstract
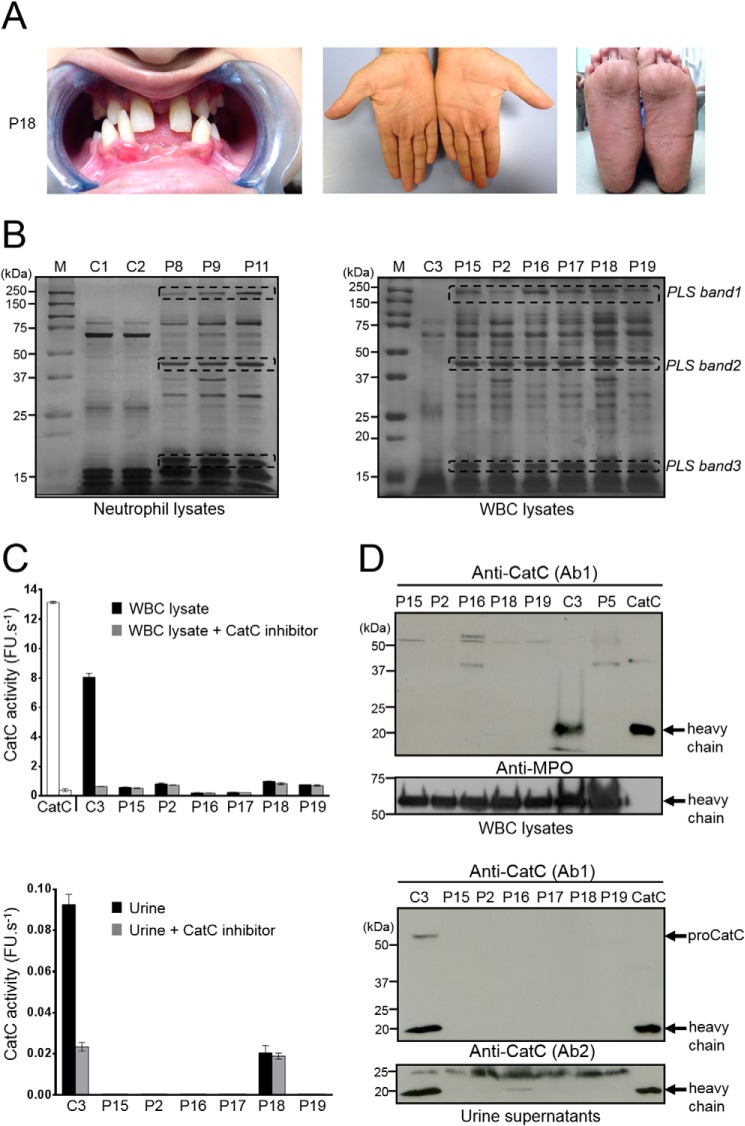
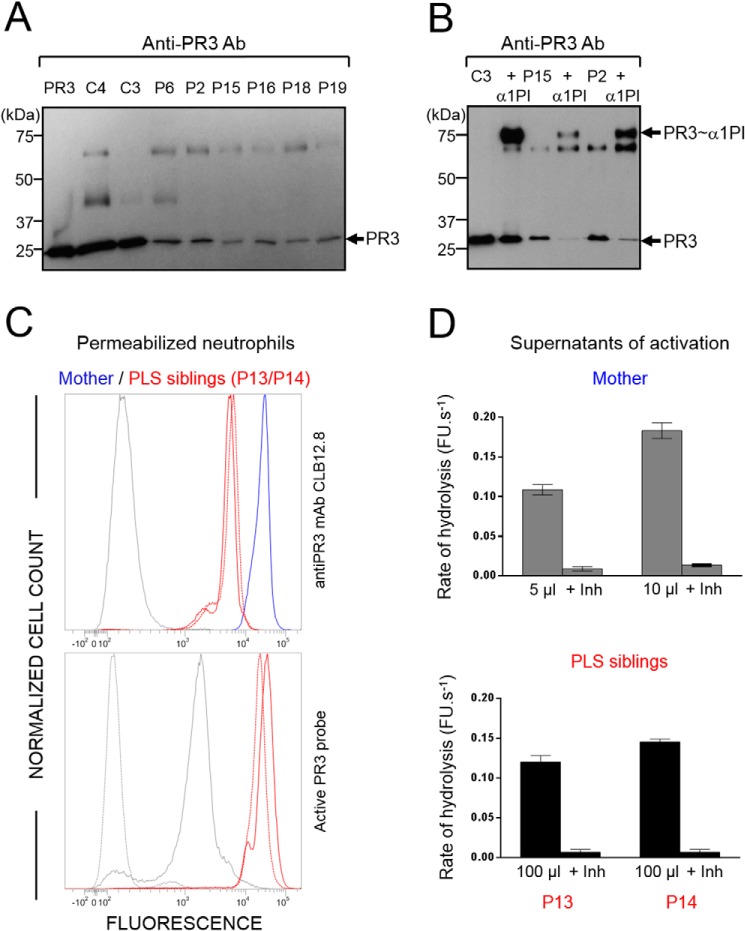
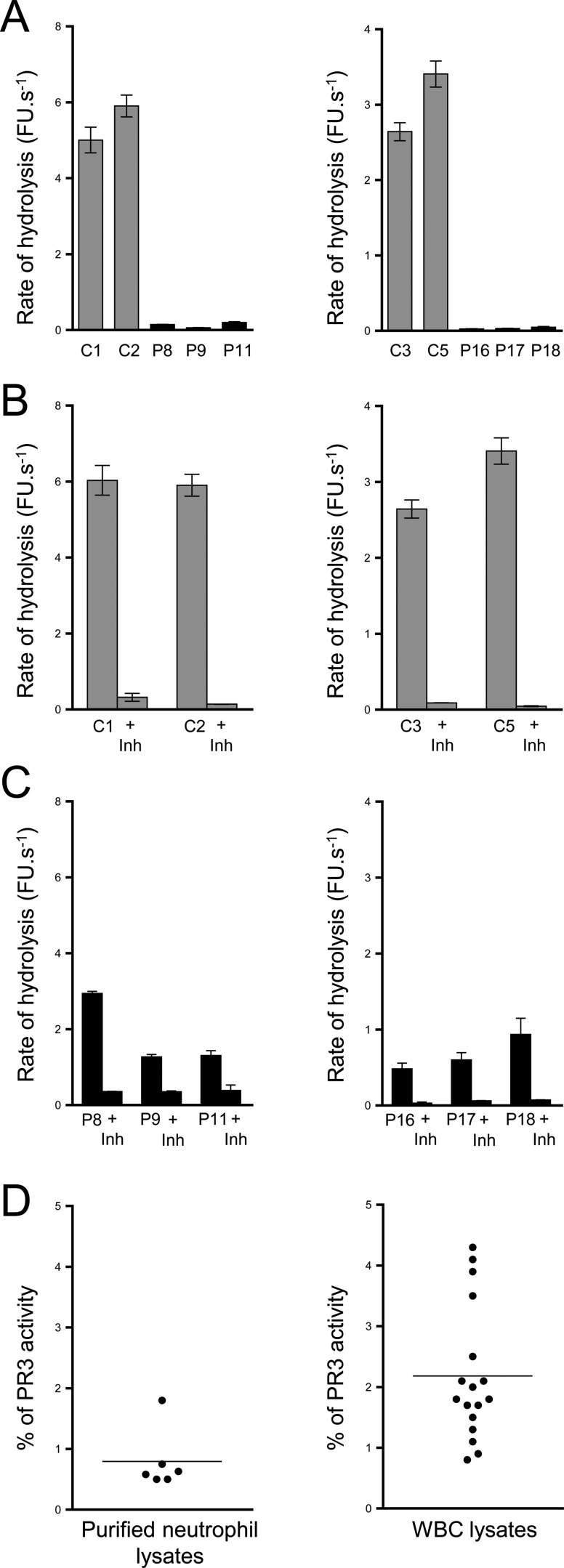
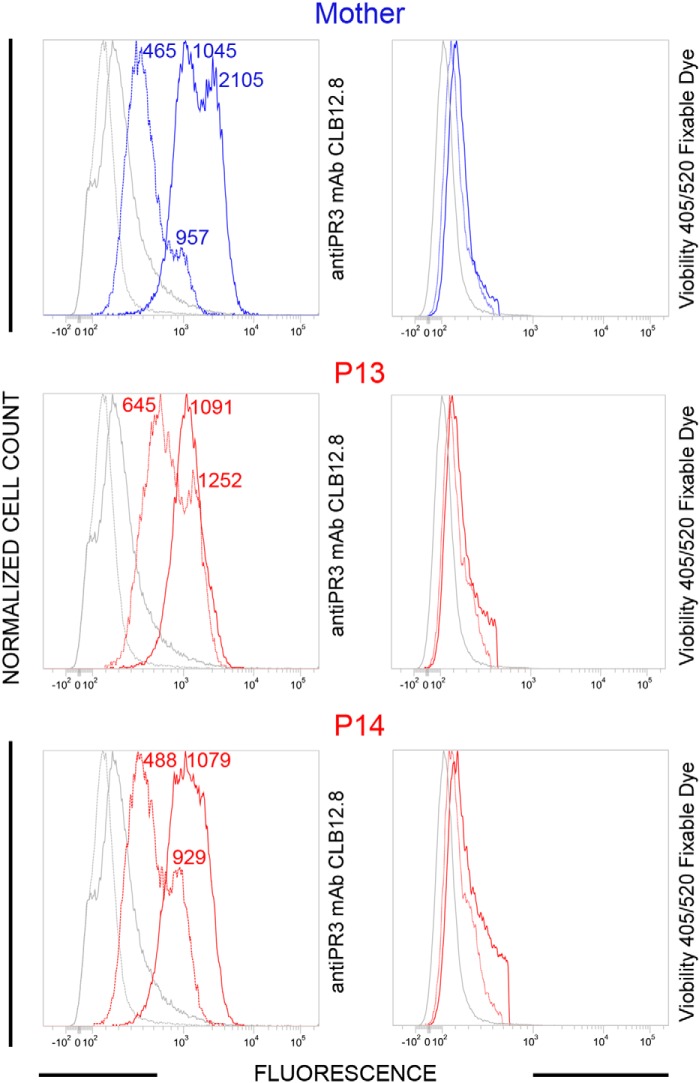
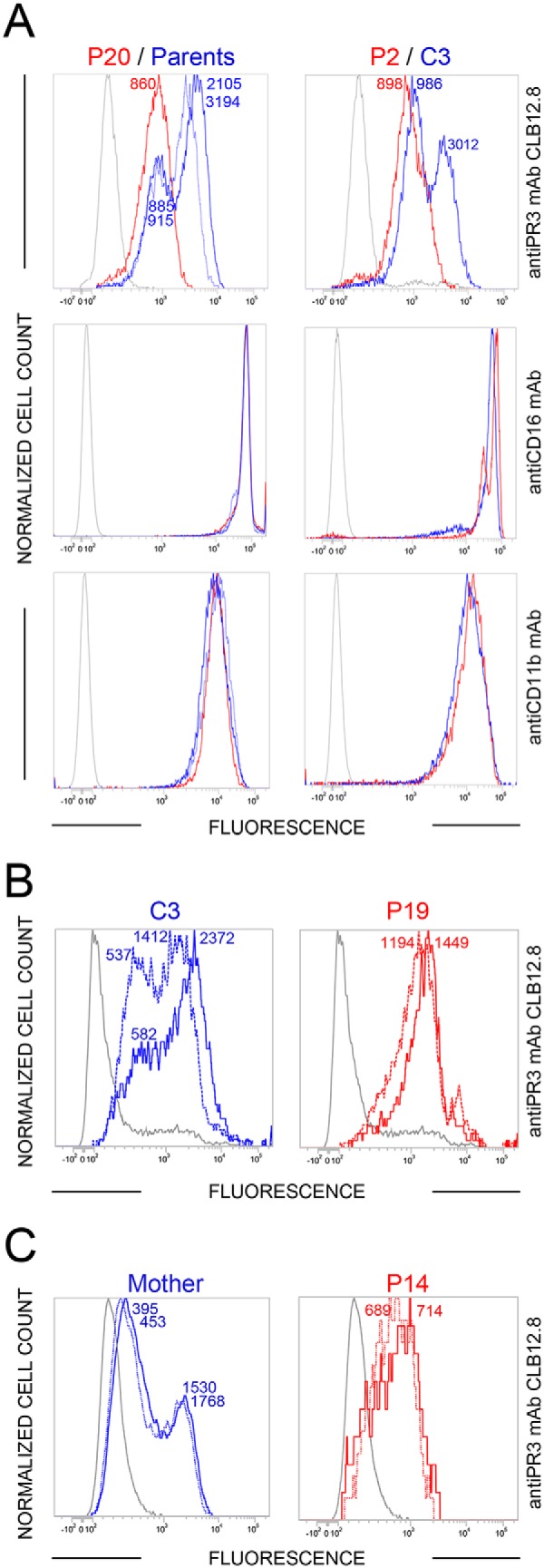
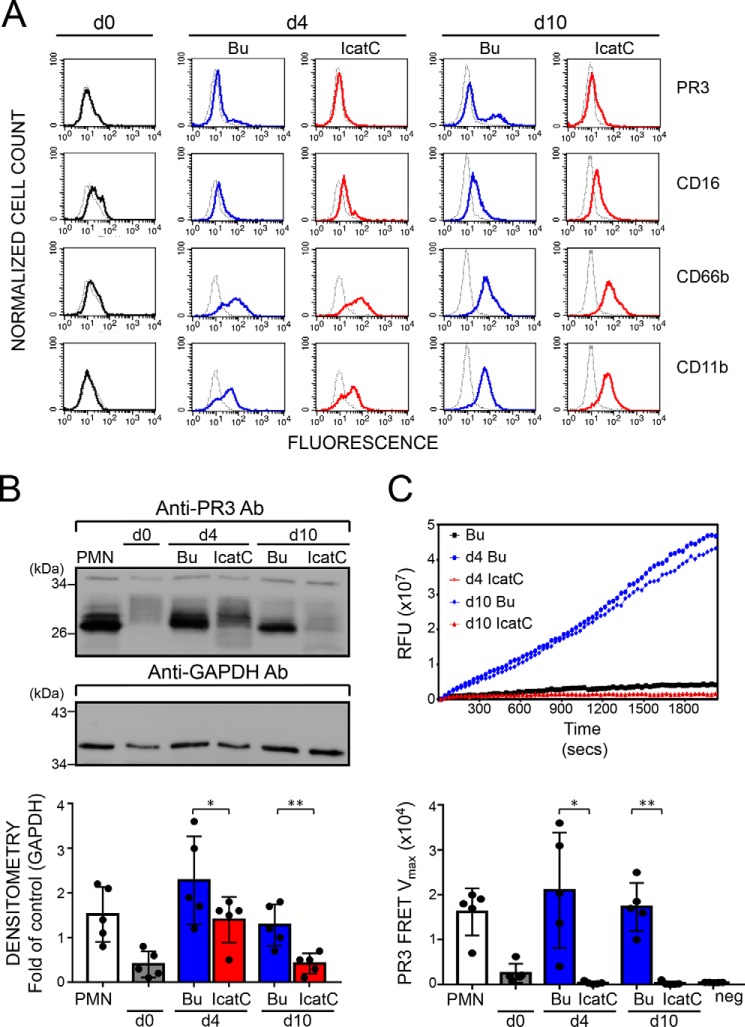
Membrane-bound proteinase 3 (PR3m) is the main target antigen of anti-neutrophil cytoplasmic autoantibodies (ANCA) in granulomatosis with polyangiitis, a systemic small-vessel vasculitis. Binding of ANCA to PR3m triggers neutrophil activation with the secretion of enzymatically active PR3 and related neutrophil serine proteases, thereby contributing to vascular damage. PR3 and related proteases are activated from pro-forms by the lysosomal cysteine protease cathepsin C (CatC) during neutrophil maturation. We hypothesized that pharmacological inhibition of CatC provides an effective measure to reduce PR3m and therefore has implications as a novel therapeutic approach in granulomatosis with polyangiitis. We first studied neutrophilic PR3 from 24 patients with Papillon-Lefèvre syndrome (PLS), a genetic form of CatC deficiency. PLS neutrophil lysates showed a largely reduced but still detectable (0.5-4%) PR3 activity when compared with healthy control cells. Despite extremely low levels of cellular PR3, the amount of constitutive PR3m expressed on the surface of quiescent neutrophils and the typical bimodal membrane distribution pattern were similar to what was observed in healthy neutrophils. However, following cell activation, there was no significant increase in the total amount of PR3m on PLS neutrophils, whereas the total amount of PR3m on healthy neutrophils was significantly increased. We then explored the effect of pharmacological CatC inhibition on PR3 stability in normal neutrophils using a potent cell-permeable CatC inhibitor and a CD34+ hematopoietic stem cell model. Human CD34+ hematopoietic stem cells were treated with the inhibitor during neutrophil differentiation over 10 days. We observed strong reductions in PR3m, cellular PR3 protein, and proteolytic PR3 activity, whereas neutrophil differentiation was not compromised.
Keywords: Papillon-Lefèvre syndrome; aminopeptidase; antigen; autoimmune disease; cathepsin C; genetic disease; granulomatosis with polyangiitis; neutrophil; protease; protease inhibitor; proteinase 3.
© 2018 Seren et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article. Responsibility for the information and views set out in this article lies entirely with the authors
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
