

Double-stranded RNA innate immune response activation from long-term adeno-associated virus vector transduction
- PMID: 29925692
- PMCID: PMC6124417
- DOI: 10.1172/jci.insight.120474
Double-stranded RNA innate immune response activation from long-term adeno-associated virus vector transduction
Abstract
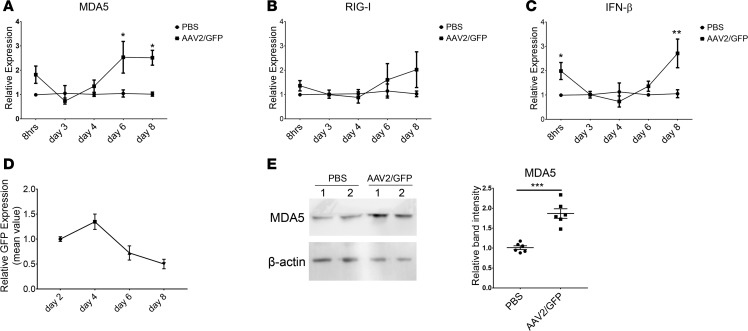
Data from clinical trials for hemophilia B using adeno-associated virus (AAV) vectors have demonstrated decreased transgenic coagulation factor IX (hFIX) expression 6-10 weeks after administration of a high vector dose. While it is likely that capsid-specific cytotoxic T lymphocytes eliminate vector-transduced hepatocytes, thereby resulting in decreased hFIX, this observation is not intuitively consistent with restored hFIX levels following prednisone application. Although the innate immune response is immediately activated following AAV vector infection via TLR pathways, no studies exist regarding the role of the innate immune response at later time points after AAV vector transduction. Herein, activation of the innate immune response in cell lines, primary human hepatocytes, and hepatocytes in a human chimeric mouse model was observed at later time points following AAV vector transduction. Mechanistic analysis demonstrated that the double-stranded RNA (dsRNA) sensor MDA5 was necessary for innate immune response activation and that transient knockdown of MDA5, or MAVS, decreased IFN-β expression while increasing transgene production in AAV-transduced cells. These results both highlight the role of the dsRNA-triggered innate immune response in therapeutic transgene expression at later time points following AAV transduction and facilitate the execution of effective strategies to block the dsRNA innate immune response in future clinical trials.
Keywords: Gene therapy; Immunology; Innate immunity; Therapeutics.
Conflict of interest statement
Figures








Similar articles
-
The role and mechanism of RLR innate immune signaling pathway in long-term AAV infection.Int Immunopharmacol. 2025 Apr 4;151:114267. doi: 10.1016/j.intimp.2025.114267. Epub 2025 Feb 20. Int Immunopharmacol. 2025. PMID: 39983417
-
Immune responses to AAV in clinical trials.Curr Gene Ther. 2007 Oct;7(5):316-24. doi: 10.2174/156652307782151425. Curr Gene Ther. 2007. PMID: 17979678 Review.
-
Pre-existing AAV capsid-specific CD8+ T cells are unable to eliminate AAV-transduced hepatocytes.Mol Ther. 2007 Apr;15(4):792-800. doi: 10.1038/sj.mt.6300090. Epub 2007 Jan 23. Mol Ther. 2007. PMID: 17245353
-
Long-term safety and efficacy following systemic administration of a self-complementary AAV vector encoding human FIX pseudotyped with serotype 5 and 8 capsid proteins.Mol Ther. 2011 May;19(5):876-85. doi: 10.1038/mt.2010.274. Epub 2011 Jan 18. Mol Ther. 2011. PMID: 21245849 Free PMC article.
-
Immune responses to AAV in clinical trials.Curr Gene Ther. 2011 Aug;11(4):321-30. doi: 10.2174/156652311796150354. Curr Gene Ther. 2011. PMID: 21557723 Review.
Cited by
-
Deep Parallel Characterization of AAV Tropism and AAV-Mediated Transcriptional Changes via Single-Cell RNA Sequencing.Front Immunol. 2021 Oct 21;12:730825. doi: 10.3389/fimmu.2021.730825. eCollection 2021. Front Immunol. 2021. PMID: 34759919 Free PMC article.
-
The curious case of AAV immunology.Mol Ther. 2025 May 7;33(5):1946-1965. doi: 10.1016/j.ymthe.2025.03.037. Epub 2025 Mar 27. Mol Ther. 2025. PMID: 40156190 Review.
-
Adeno-Associated Viruses (AAV) and Host Immunity - A Race Between the Hare and the Hedgehog.Front Immunol. 2021 Oct 29;12:753467. doi: 10.3389/fimmu.2021.753467. eCollection 2021. Front Immunol. 2021. PMID: 34777364 Free PMC article. Review.
-
Immune Responses to Viral Gene Therapy Vectors.Mol Ther. 2020 Mar 4;28(3):709-722. doi: 10.1016/j.ymthe.2020.01.001. Epub 2020 Jan 10. Mol Ther. 2020. PMID: 31968213 Free PMC article. Review.
-
Immunogenicity assessment of AAV-based gene therapies: An IQ consortium industry white paper.Mol Ther Methods Clin Dev. 2022 Aug 2;26:471-494. doi: 10.1016/j.omtm.2022.07.018. eCollection 2022 Sep 8. Mol Ther Methods Clin Dev. 2022. PMID: 36092368 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
