

The role of calretinin-expressing granule cells in olfactory bulb functions and odor behavior
- PMID: 29925844
- PMCID: PMC6010413
- DOI: 10.1038/s41598-018-27692-8
The role of calretinin-expressing granule cells in olfactory bulb functions and odor behavior
Abstract
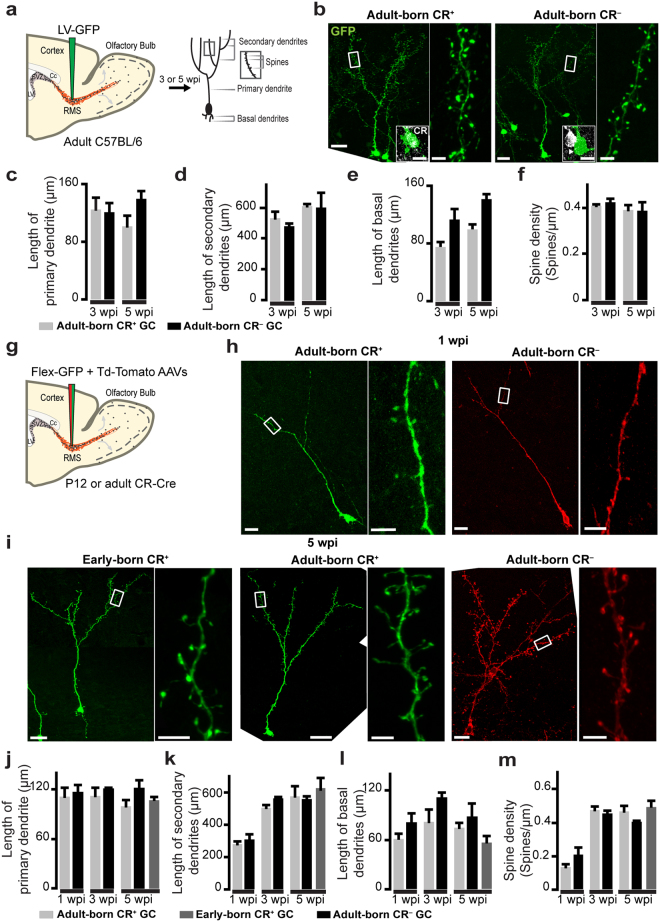
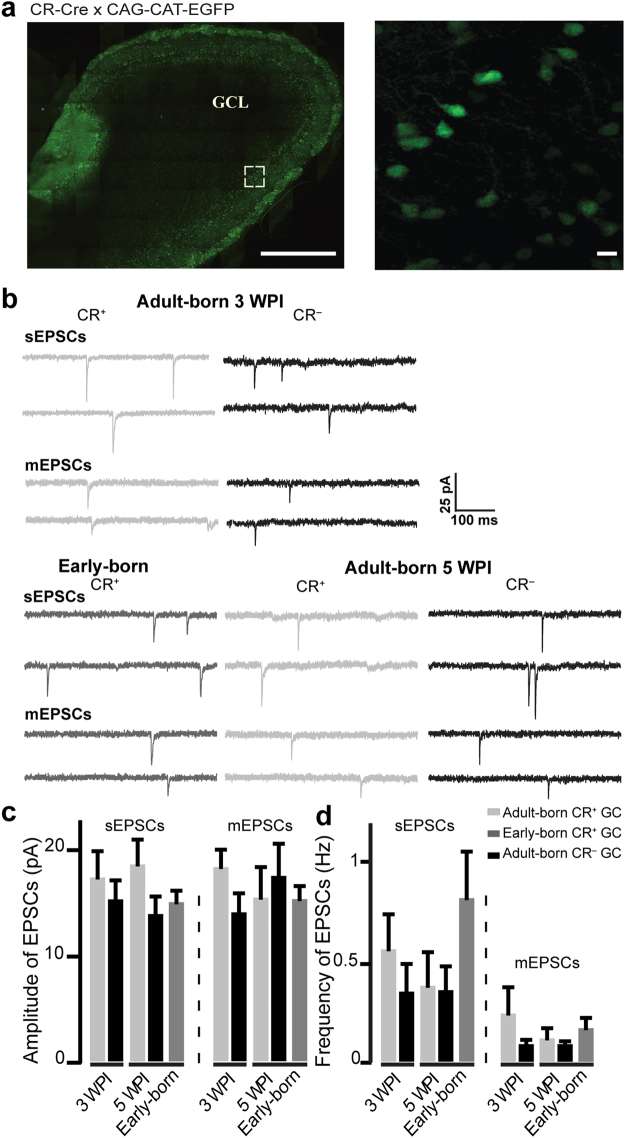
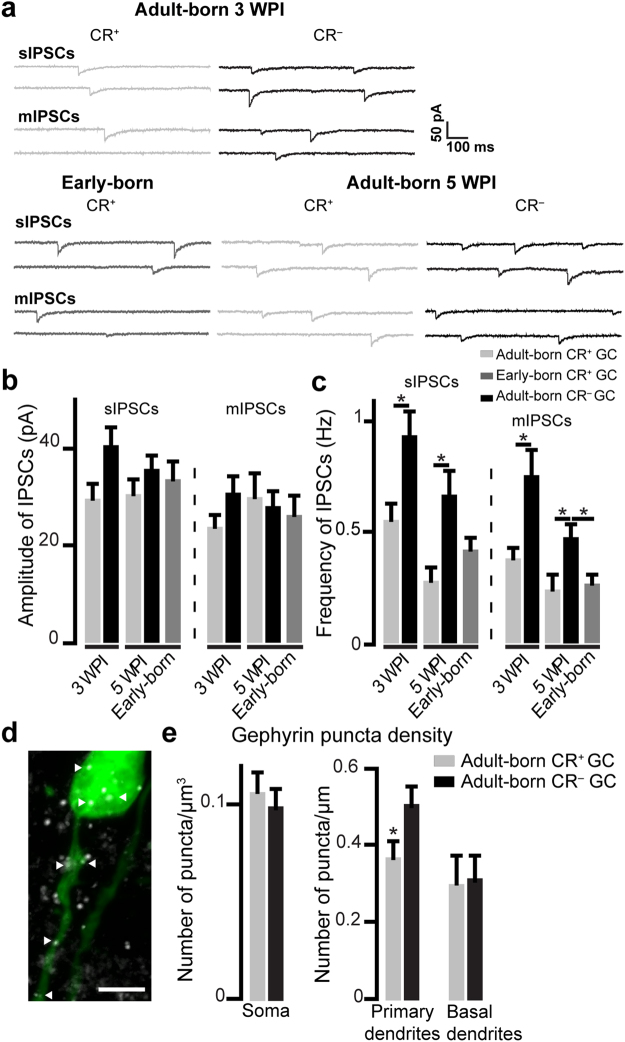
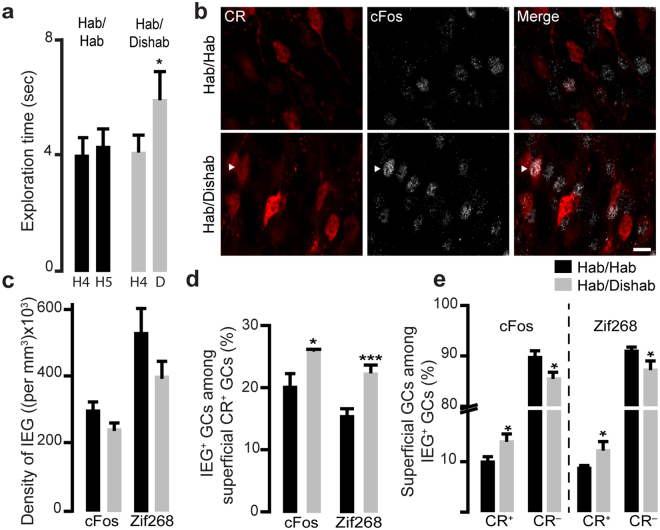
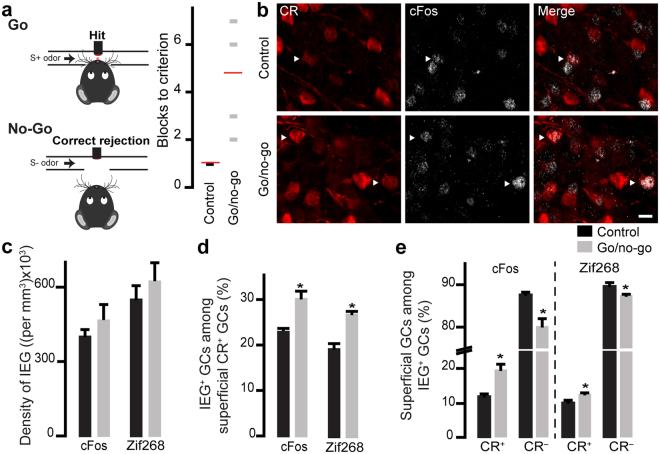
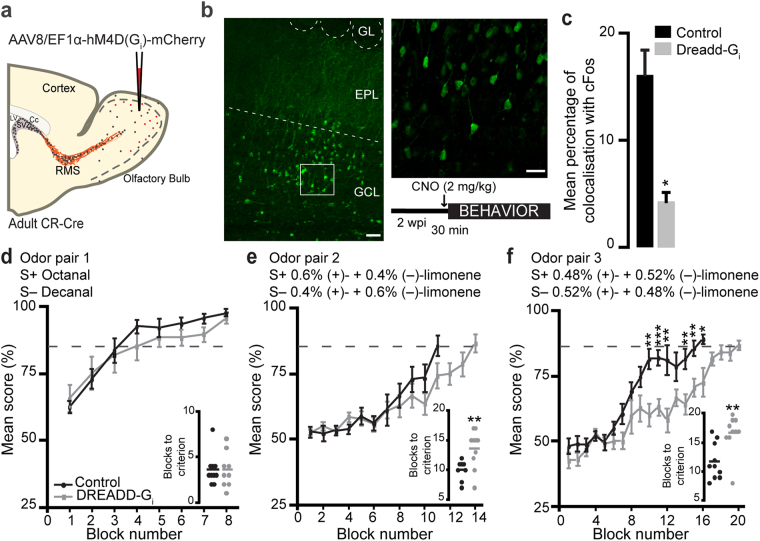
The adult mouse olfactory bulb is continuously supplied with new neurons that mostly differentiate into granule cells (GCs). Different subtypes of adult-born GCs have been identified, but their maturational profiles and their roles in bulbar network functioning and odor behavior remain elusive. It is also not known whether the same subpopulations of GCs born during early postnatal life (early-born) or during adulthood (adult-born) differ in their morpho-functional properties. Here, we show that adult-born calretinin-expressing (CR+) and non-expressing (CR-) GCs, as well as early-born CR+ GCs, display distinct inhibitory inputs but indistinguishable excitatory inputs and similar morphological characteristics. The frequencies of inhibitory post-synaptic currents were lower in early-born and adult-born CR+ GCs than in adult-born CR- neurons. These findings were corroborated by the reduced density of gephyrin+ puncta on CR+ GCs. CR+ GCs displayed a higher level of activation following olfactory tasks based on odor discrimination, as determined by an immediate early gene expression analysis. Pharmacogenetic inhibition of CR+ GCs diminished the ability of the mice to discriminate complex odor mixtures. Altogether, our results indicate that distinct inhibitory inputs are received by adult-born CR+ and CR- GCs, that early- and adult-born CR+ neurons have similar morpho-functional properties, and that CR+ GCs are involved in complex odor discrimination tasks.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
