

A naturally occurring antiviral ribonucleotide encoded by the human genome
- PMID: 29925952
- PMCID: PMC6026066
- DOI: 10.1038/s41586-018-0238-4
A naturally occurring antiviral ribonucleotide encoded by the human genome
Erratum in
-
Publisher Correction: A naturally occurring antiviral ribonucleotide encoded by the human genome.Nature. 2018 Oct;562(7725):E3. doi: 10.1038/s41586-018-0355-0. Nature. 2018. PMID: 29980769
-
Author Correction: A naturally occurring antiviral ribonucleotide encoded by the human genome.Nature. 2020 Jul;583(7814):E15. doi: 10.1038/s41586-020-2322-9. Nature. 2020. PMID: 32541969
Abstract
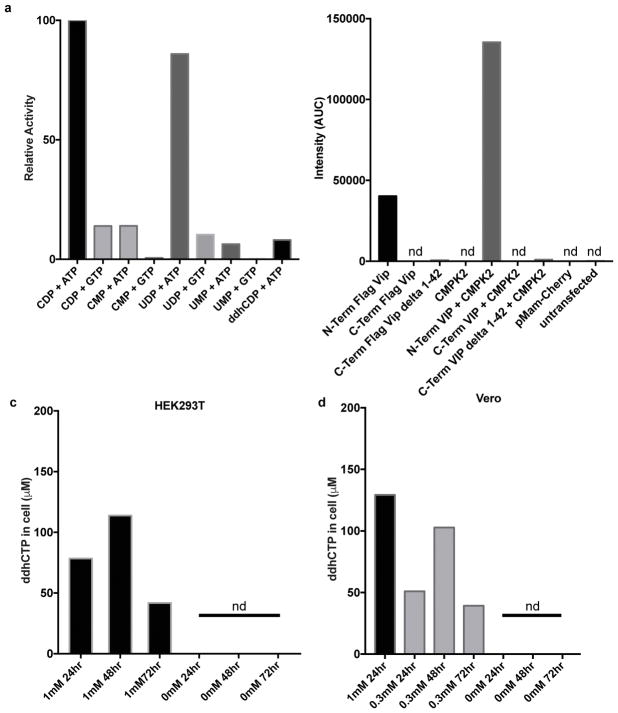
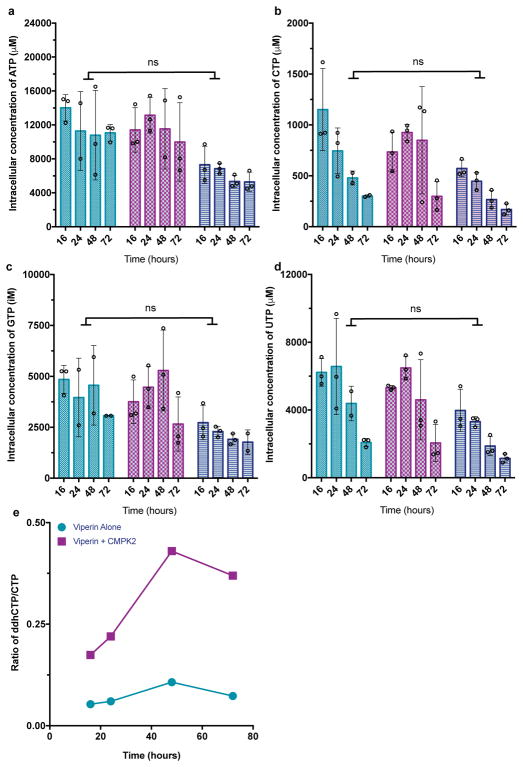
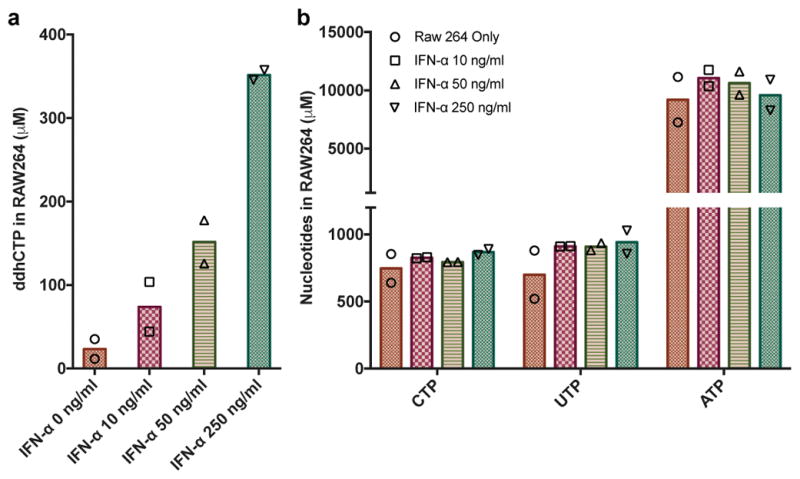
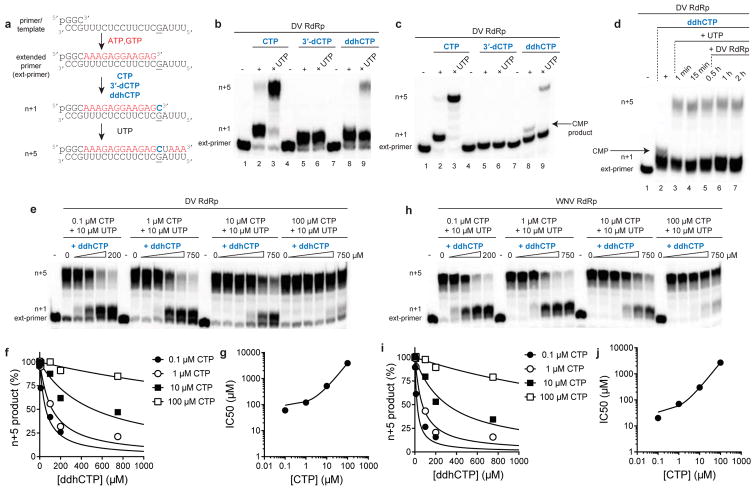
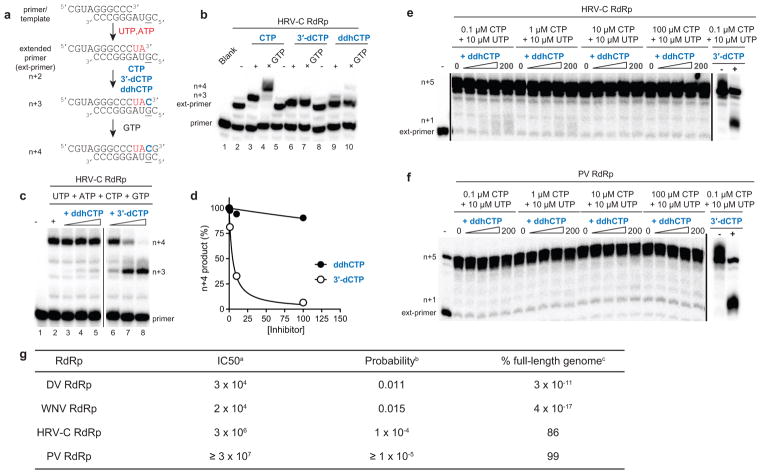
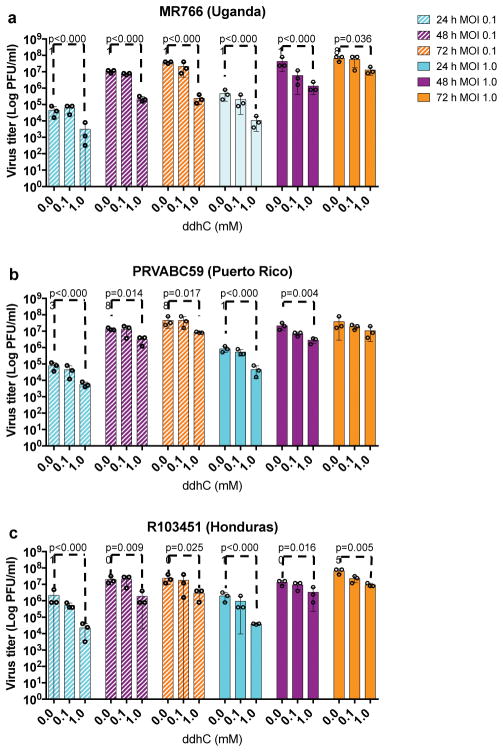
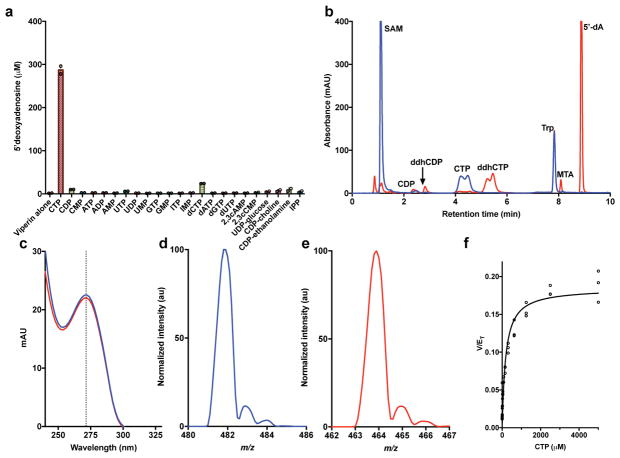
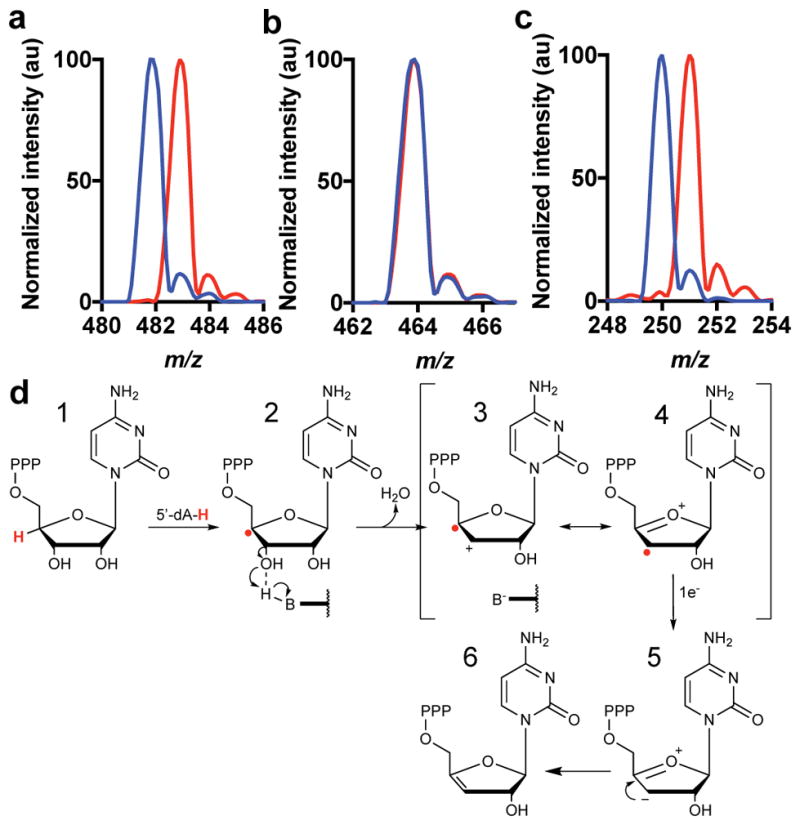
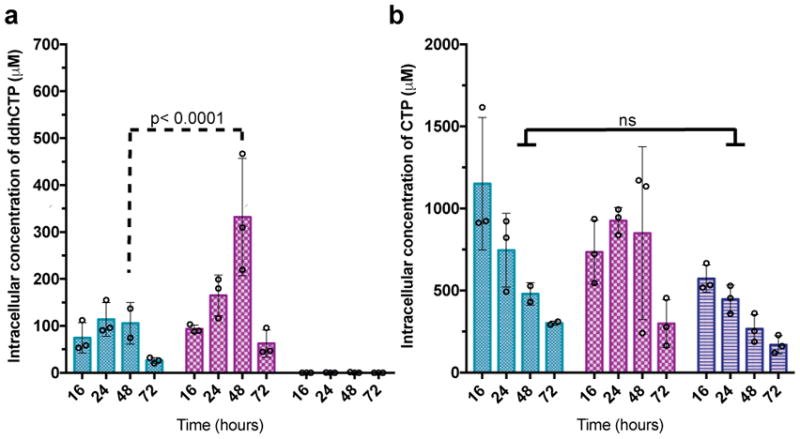
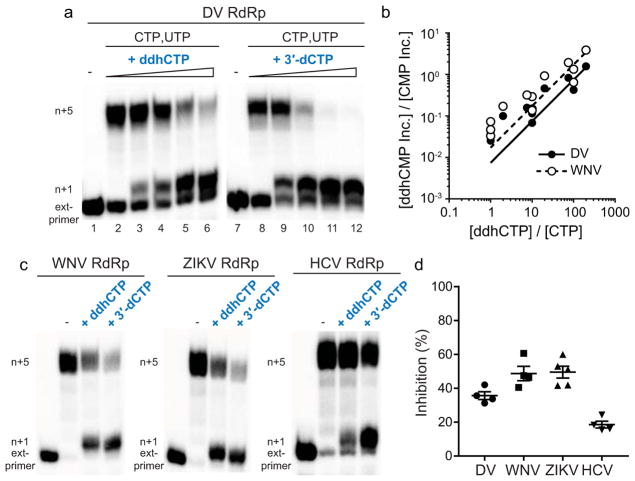
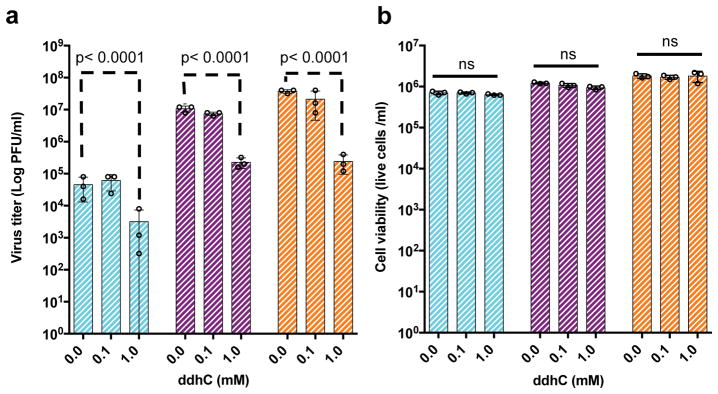
Viral infections continue to represent major challenges to public health, and an enhanced mechanistic understanding of the processes that contribute to viral life cycles is necessary for the development of new therapeutic strategies 1 . Viperin, a member of the radical S-adenosyl-L-methionine (SAM) superfamily of enzymes, is an interferon-inducible protein implicated in the inhibition of replication of a broad range of RNA and DNA viruses, including dengue virus, West Nile virus, hepatitis C virus, influenza A virus, rabies virus 2 and HIV3,4. Viperin has been suggested to elicit these broad antiviral activities through interactions with a large number of functionally unrelated host and viral proteins3,4. Here we demonstrate that viperin catalyses the conversion of cytidine triphosphate (CTP) to 3'-deoxy-3',4'-didehydro-CTP (ddhCTP), a previously undescribed biologically relevant molecule, via a SAM-dependent radical mechanism. We show that mammalian cells expressing viperin and macrophages stimulated with IFNα produce substantial quantities of ddhCTP. We also establish that ddhCTP acts as a chain terminator for the RNA-dependent RNA polymerases from multiple members of the Flavivirus genus, and show that ddhCTP directly inhibits replication of Zika virus in vivo. These findings suggest a partially unifying mechanism for the broad antiviral effects of viperin that is based on the intrinsic enzymatic properties of the protein and involves the generation of a naturally occurring replication-chain terminator encoded by mammalian genomes.
Conflict of interest statement
A.S.G., T.L.G., J.J.A., C.E.C., and S.C.A. are co-inventors on a U.S. provisional patent application (No. 62/548,425; filed by S.C.A) that incorporates discoveries described in this manuscript.
Figures
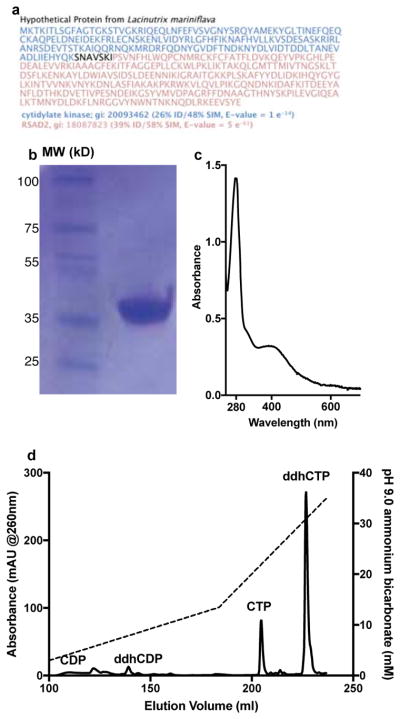
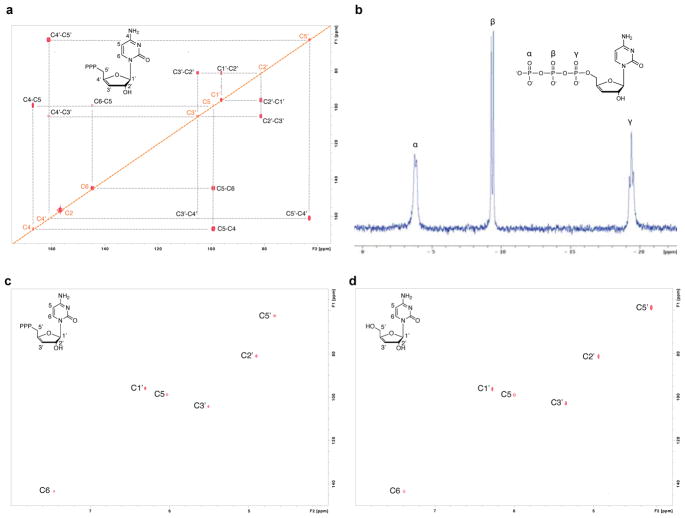
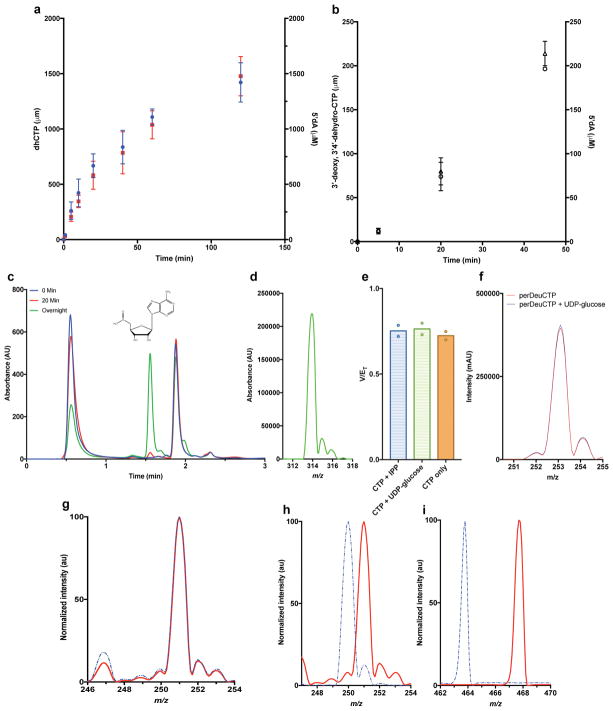
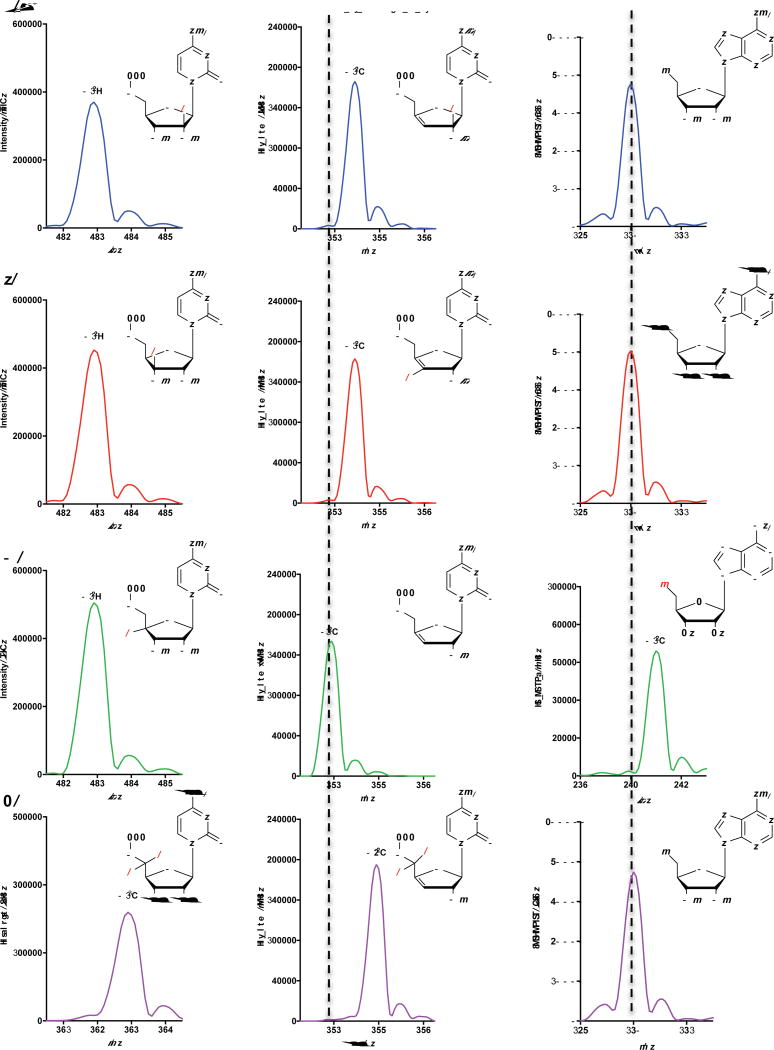
Comment in
-
Viperin breaks viral chains.Nat Rev Immunol. 2018 Aug;18(8):480-481. doi: 10.1038/s41577-018-0035-1. Nat Rev Immunol. 2018. PMID: 29950609 No abstract available.
-
Viperin Poisons Viral Replication.Cell Host Microbe. 2018 Aug 8;24(2):181-183. doi: 10.1016/j.chom.2018.07.014. Cell Host Microbe. 2018. PMID: 30092190
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
