

Synaptic aging disrupts synaptic morphology and function in cerebellar Purkinje cells
- PMID: 29926829
- PMCID: PMC6022458
- DOI: 10.4103/1673-5374.233445
Synaptic aging disrupts synaptic morphology and function in cerebellar Purkinje cells
Abstract
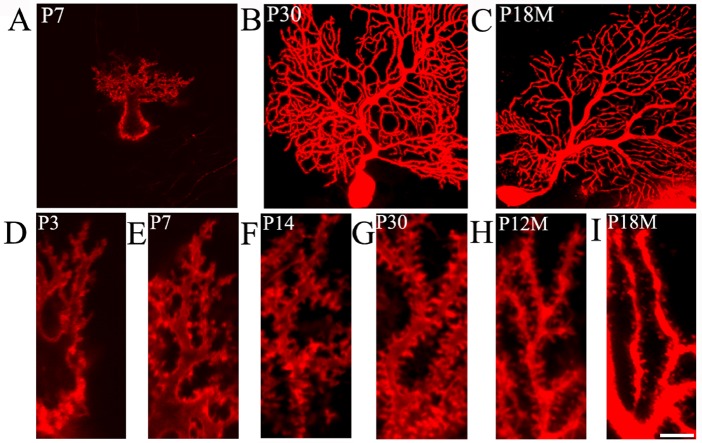
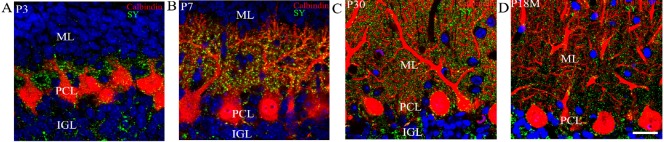
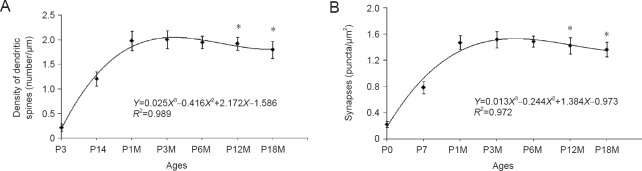
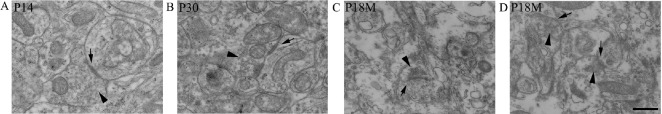
Synapses are key structures in neural networks, and are involved in learning and memory in the central nervous system. Investigating synaptogenesis and synaptic aging is important in understanding neural development and neural degeneration in diseases such as Alzheimer disease and Parkinson's disease. Our previous study found that synaptogenesis and synaptic maturation were harmonized with brain development and maturation. However, synaptic damage and loss in the aging cerebellum are not well understood. This study was designed to investigate the occurrence of synaptic aging in the cerebellum by observing the ultrastructural changes of dendritic spines and synapses in cerebellar Purkinje cells of aging mice. Immunocytochemistry, DiI diolistic assays, and transmission electron microscopy were used to visualize the morphological characteristics of synaptic buttons, dendritic spines and synapses of Purkinje cells in mice at various ages. With synaptic aging in the cerebellum, dendritic spines and synaptic buttons were lost, and the synaptic ultrastructure was altered, including a reduction in the number of synaptic vesicles and mitochondria in presynaptic termini and smaller thin specialized zones in pre- and post-synaptic membranes. These findings confirm that synaptic morphology and function is disrupted in aging synapses, which may be an important pathological cause of neurodegenerative diseases.
Keywords: Purkinje cells; aging; cerebellum; degenerative disease; dendritic spine; mice; nerve regeneration; neural regeneration; neurodegenerative diseases; synapse; synaptic ultrastructure; synaptogenesis.
Conflict of interest statement
The authors declare no competing financial interests
Figures
Similar articles
-
Serial changes in granuloprival cerebellar cultures after transplantation with granule cells and glia: a timed ultrastructural study.Neuroscience. 1997 Apr;77(3):695-711. doi: 10.1016/s0306-4522(96)00546-5. Neuroscience. 1997. PMID: 9070746
-
Synaptogenesis in the developing mouse visual cortex.Brain Res Bull. 2010 Jan 15;81(1):107-13. doi: 10.1016/j.brainresbull.2009.08.028. Brain Res Bull. 2010. PMID: 19751806
-
Morphological development of dendritic spines on rat cerebellar Purkinje cells.Int J Dev Neurosci. 2011 Aug;29(5):515-20. doi: 10.1016/j.ijdevneu.2011.04.005. Epub 2011 Apr 23. Int J Dev Neurosci. 2011. PMID: 21549828
-
Development and fine structure of murine Purkinje cells in dissociated cerebellar cultures: dendritic differentiation, synaptic maturation, and formation of cell-class specific features.Anat Embryol (Berl). 1998 Jan;197(1):31-50. doi: 10.1007/s004290050118. Anat Embryol (Berl). 1998. PMID: 9462857 Review.
-
Synaptic Plasticity, Dementia and Alzheimer Disease.CNS Neurol Disord Drug Targets. 2017;16(3):220-233. doi: 10.2174/1871527316666170113120853. CNS Neurol Disord Drug Targets. 2017. PMID: 28088900 Review.
Cited by
-
Neurexin and neuroligins jointly regulate synaptic degeneration at the Drosophila neuromuscular junction based on TEM studies.Front Cell Neurosci. 2023 Nov 1;17:1257347. doi: 10.3389/fncel.2023.1257347. eCollection 2023. Front Cell Neurosci. 2023. PMID: 38026694 Free PMC article.
-
Imaging Synaptic Density: The Next Holy Grail of Neuroscience?Front Neurosci. 2022 Mar 25;16:796129. doi: 10.3389/fnins.2022.796129. eCollection 2022. Front Neurosci. 2022. PMID: 35401097 Free PMC article. Review.
-
Celsr3 is required for Purkinje cell maturation and regulates cerebellar postsynaptic plasticity.iScience. 2021 Jul 1;24(7):102812. doi: 10.1016/j.isci.2021.102812. eCollection 2021 Jul 23. iScience. 2021. PMID: 34308297 Free PMC article.
-
Aging, Neurodegenerative Disorders, and Cerebellum.Int J Mol Sci. 2024 Jan 13;25(2):1018. doi: 10.3390/ijms25021018. Int J Mol Sci. 2024. PMID: 38256091 Free PMC article. Review.
-
Synaptic vesicle-omics in mice captures signatures of aging and synucleinopathy.Nat Commun. 2025 May 1;16(1):4079. doi: 10.1038/s41467-025-59441-7. Nat Commun. 2025. PMID: 40312501 Free PMC article.
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
