

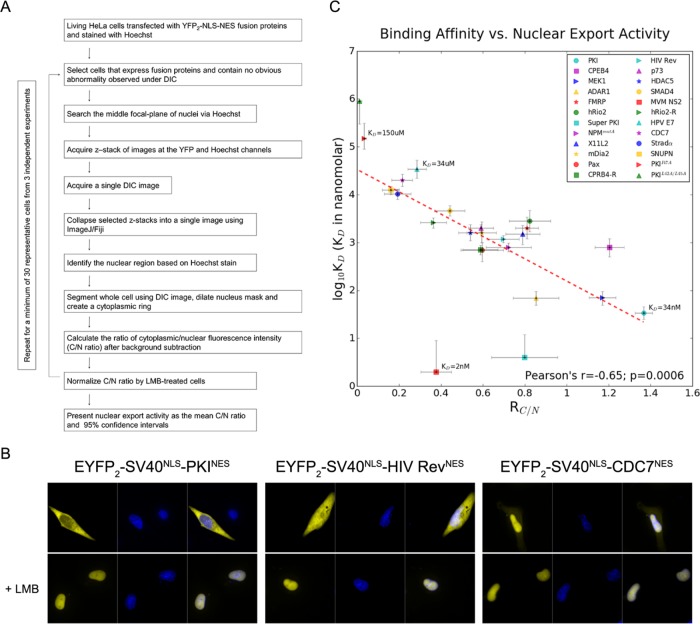
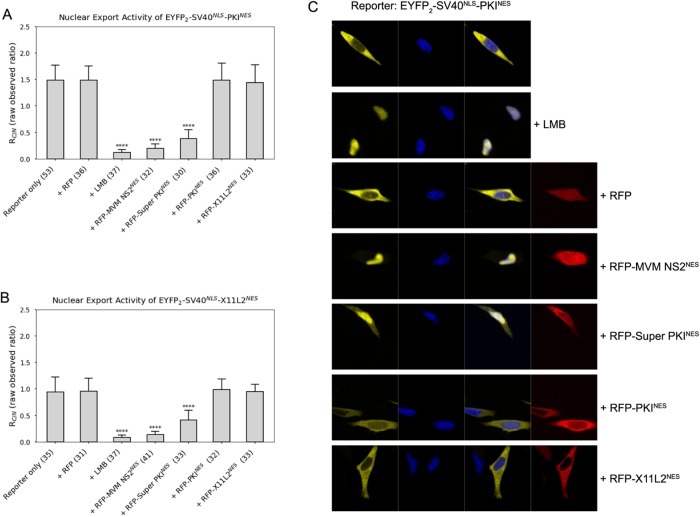
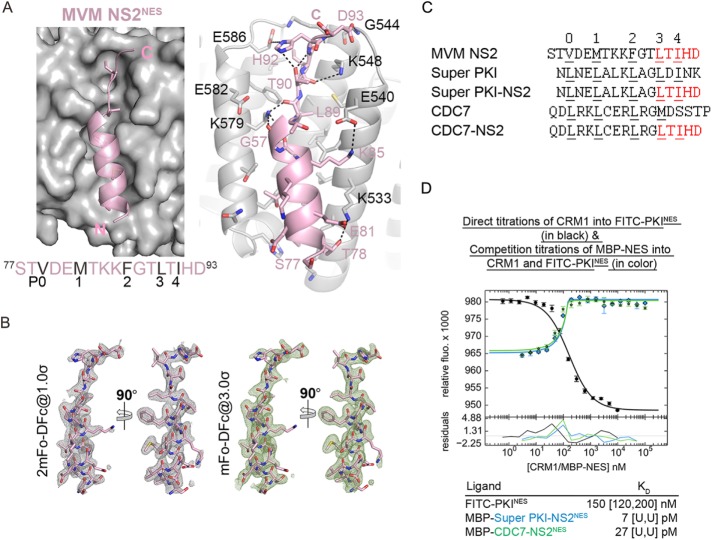
Correlation of CRM1-NES affinity with nuclear export activity
- PMID: 29927350
- PMCID: PMC6232958
- DOI: 10.1091/mbc.E18-02-0096
Correlation of CRM1-NES affinity with nuclear export activity
Abstract
CRM1 (Exportin1/XPO1) exports hundreds of broadly functioning protein cargoes out of the cell nucleus by binding to their classical nuclear export signals (NESs). The 8- to 15-amino-acid-long NESs contain four to five hydrophobic residues and are highly diverse in both sequence and CRM1-bound structure. Here we examine the relationship between nuclear export activities of 24 different NES peptides in cells and their CRM1-NES affinities. We found that binding affinity and nuclear export activity are linearly correlated for NESs with dissociation constants ( Kds) between tens of nanomolar to tens of micromolar. NESs with Kds outside this range have significantly reduced nuclear export activities. These include two unusually tight-binding peptides, one from the nonstructural protein 2 of murine minute virus (MVM NS2) and the other a mutant of the protein kinase A inhibitor (PKI) NES. The crystal structure of CRM1-bound MVM NS2NES suggests that extraordinarily tight CRM1 binding arises from intramolecular contacts within the NES that likely stabilizes the CRM1-bound conformation in free peptides. This mechanistic understanding led to the design of two novel peptide inhibitors that bind CRM1 with picomolar affinity.
Figures
Similar articles
-
Nuclear export receptor CRM1 recognizes diverse conformations in nuclear export signals.Elife. 2017 Mar 10;6:e23961. doi: 10.7554/eLife.23961. Elife. 2017. PMID: 28282025 Free PMC article.
-
Binding Affinity Measurement of Nuclear Export Signal Peptides to Their Exporter CRM1.Methods Mol Biol. 2022;2502:245-256. doi: 10.1007/978-1-0716-2337-4_16. Methods Mol Biol. 2022. PMID: 35412243
-
Structural determinants of nuclear export signal orientation in binding to exportin CRM1.Elife. 2015 Sep 8;4:e10034. doi: 10.7554/eLife.10034. Elife. 2015. PMID: 26349033 Free PMC article.
-
Inhibition of CRM1-dependent nuclear export sensitizes malignant cells to cytotoxic and targeted agents.Semin Cancer Biol. 2014 Aug;27:62-73. doi: 10.1016/j.semcancer.2014.03.001. Epub 2014 Mar 12. Semin Cancer Biol. 2014. PMID: 24631834 Free PMC article. Review.
-
Leucine-rich nuclear-export signals: born to be weak.Trends Cell Biol. 2005 Mar;15(3):121-4. doi: 10.1016/j.tcb.2005.01.005. Trends Cell Biol. 2005. PMID: 15752974 Review.
Cited by
-
Exportins can inhibit major mitotic assembly events in vitro: membrane fusion, nuclear pore formation, and spindle assembly.Nucleus. 2020 Dec;11(1):178-193. doi: 10.1080/19491034.2020.1798093. Nucleus. 2020. PMID: 32762441 Free PMC article.
-
Identification of Novel Covalent XPO1 Inhibitors Based on a Hybrid Virtual Screening Strategy.Molecules. 2022 Apr 14;27(8):2543. doi: 10.3390/molecules27082543. Molecules. 2022. PMID: 35458742 Free PMC article.
-
Cancer Therapy with Nanoparticle-Medicated Intracellular Expression of Peptide CRM1-Inhibitor.Int J Nanomedicine. 2021 Apr 14;16:2833-2847. doi: 10.2147/IJN.S266398. eCollection 2021. Int J Nanomedicine. 2021. PMID: 33883894 Free PMC article.
-
Targeting the chromatin binding of exportin-1 disrupts NFAT and T cell activation.Nat Chem Biol. 2024 Oct;20(10):1260-1271. doi: 10.1038/s41589-024-01586-5. Epub 2024 Mar 25. Nat Chem Biol. 2024. PMID: 38528120
-
Retroviral adapters hijack the RNA helicase UPF1 in a CRM1/XPO1-dependent manner and reveal proviral roles of UPF1.Nucleic Acids Res. 2025 May 10;53(9):gkaf434. doi: 10.1093/nar/gkaf434. Nucleic Acids Res. 2025. PMID: 40396490 Free PMC article.
References
-
- Conti E, Uy M, Leighton L, Blobel G, Kuriyan J. (1998). Crystallographic analysis of the recognition of a nuclear localization signal by the nuclear import factor karyopherin α. Cell , 193–204. - PubMed
-
- Fanara P, Hodel MR, Corbett AH, Hodel AE. (2000). Quantitative analysis of nuclear localization signal (NLS)-importin α interaction through fluorescence ddepolarization. J Biol Chem , 21218–21223. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
