

Egr-1: A Candidate Transcription Factor Involved in Molecular Processes Underlying Time-Memory
- PMID: 29928241
- PMCID: PMC5997935
- DOI: 10.3389/fpsyg.2018.00865
Egr-1: A Candidate Transcription Factor Involved in Molecular Processes Underlying Time-Memory
Abstract
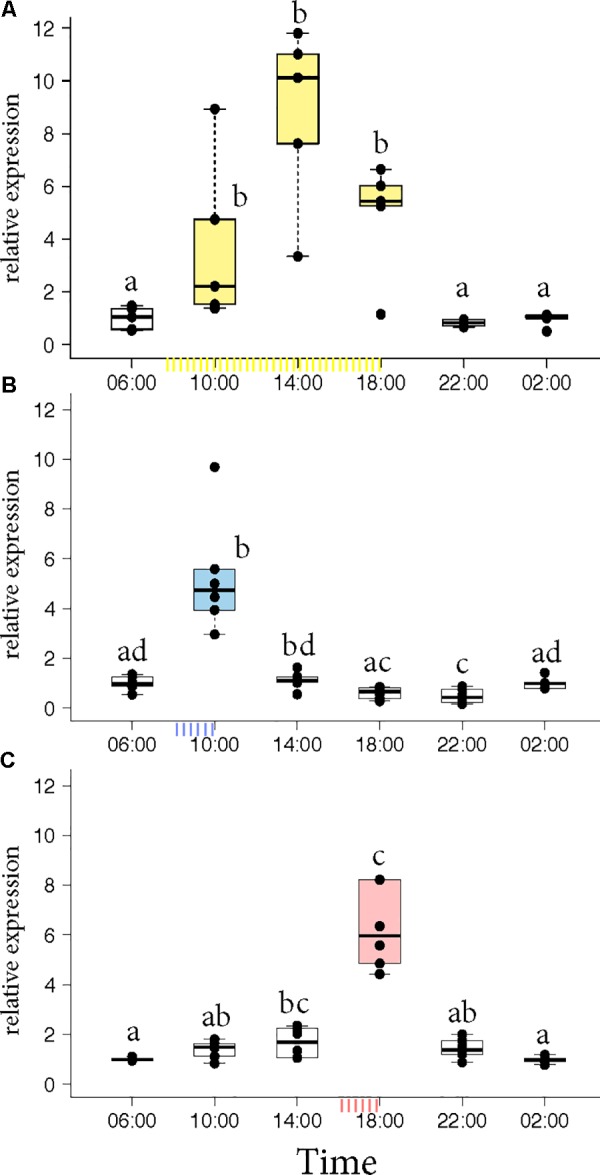
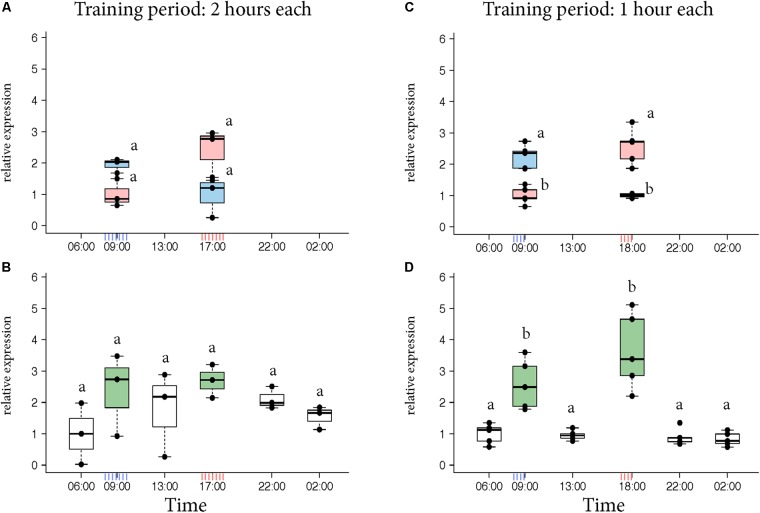
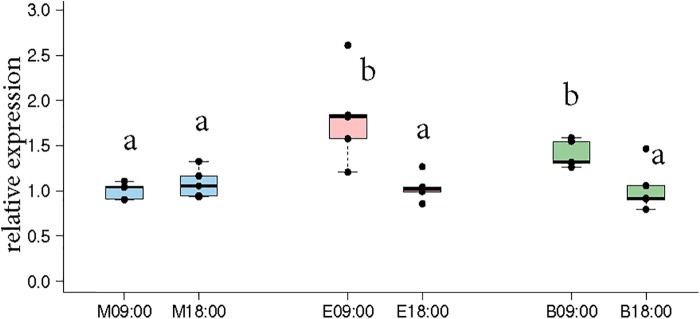
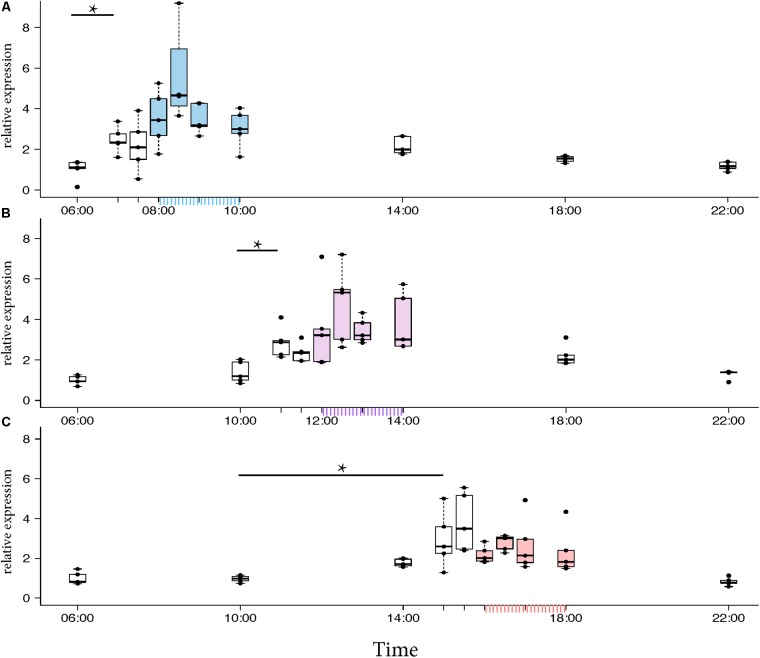
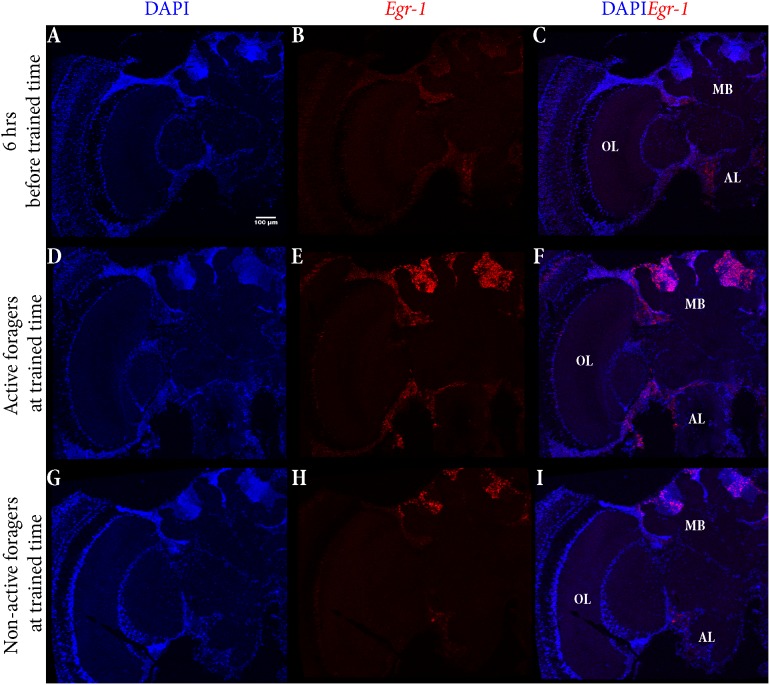
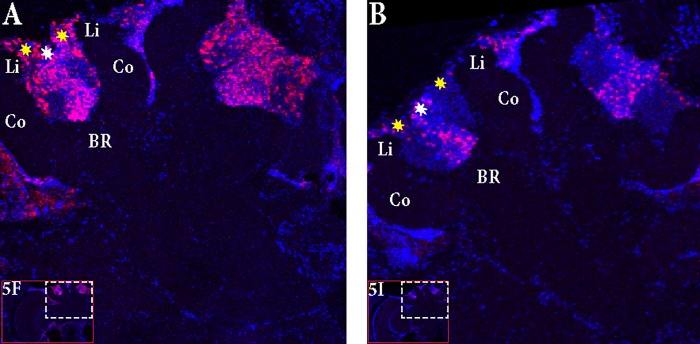
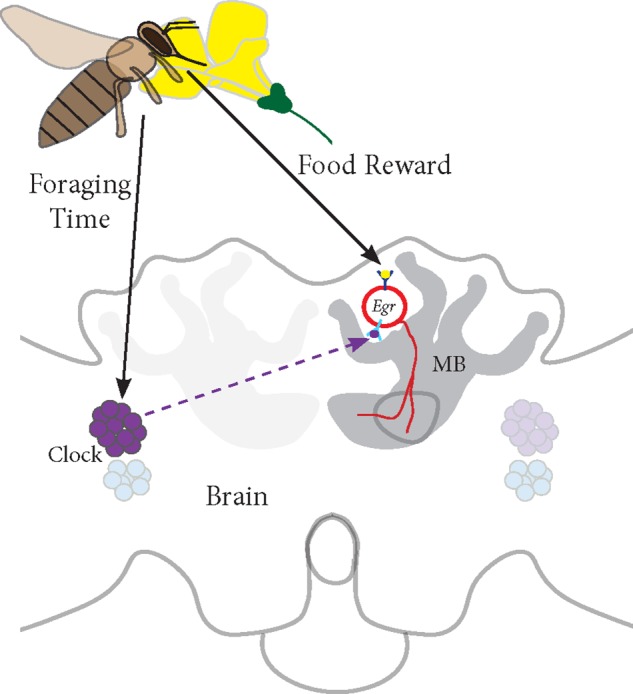
In honey bees, continuous foraging is accompanied by a sustained up-regulation of the immediate early gene Egr-1 (early growth response protein-1) and candidate downstream genes involved in learning and memory. Here, we present a series of feeder training experiments indicating that Egr-1 expression is highly correlated with the time and duration of training even in the absence of the food reward. Foragers that were trained to visit a feeder over the whole day and then collected on a day without food presentation showed Egr-1 up-regulation over the whole day with a peak expression around 14:00. When exposed to a time-restricted feeder presentation, either 2 h in the morning or 2 h in the evening, Egr-1 expression in the brain was up-regulated only during the hours of training. Foragers that visited a feeder in the morning as well as in the evening showed two peaks of Egr-1 expression. Finally, when we prevented time-trained foragers from leaving the colony using artificial rain, Egr-1 expression in the brains was still slightly but significantly up-regulated around the time of feeder training. In situ hybridization studies showed that active foraging and time-training induced Egr-1 up-regulation occurred in the same brain areas, preferentially the small Kenyon cells of the mushroom bodies and the antennal and optic lobes. Based on these findings we propose that foraging induced Egr-1 expression can get regulated by the circadian clock after time-training over several days and Egr-1 is a candidate transcription factor involved in molecular processes underlying time-memory.
Keywords: Egr-1; anticipation; honey bee foraging; small Kenyon cells; time-memory.
Figures
References
-
- Beier W. (1968). Beeinflussung der inneren Uhr der Bienen durch Phasenverschiebung des Licht–Dunkel–Zeitgebers. Zeitschrift Bienenforschung 9 356–378.
-
- Beling I. (1929). Über das Zeitgedächtnis der Bienen. Z. Vergl. Physiol. 9 259–338. 10.1007/BF00340159 - DOI
-
- Benjamini Y., Hochberg Y. (1995). Controlling the false discovery rate: a practical and powerful approach to multiple testing. J. R. Stat. Soc. Series B 57 289–300. 10.2307/2346101 - DOI
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials