

A parthenogenetic quasi-program causes teratoma-like tumors during aging in wild-type C. elegans
- PMID: 29928508
- PMCID: PMC5998035
- DOI: 10.1038/s41514-018-0025-3
A parthenogenetic quasi-program causes teratoma-like tumors during aging in wild-type C. elegans
Abstract
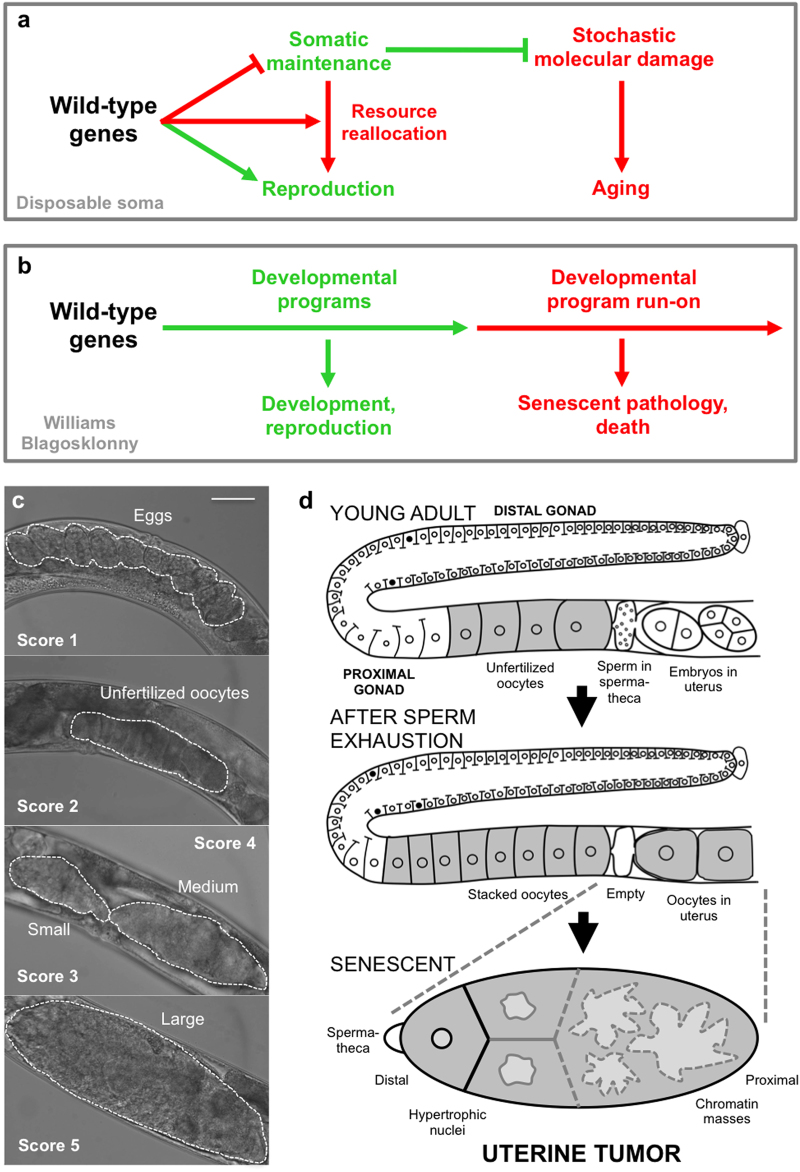
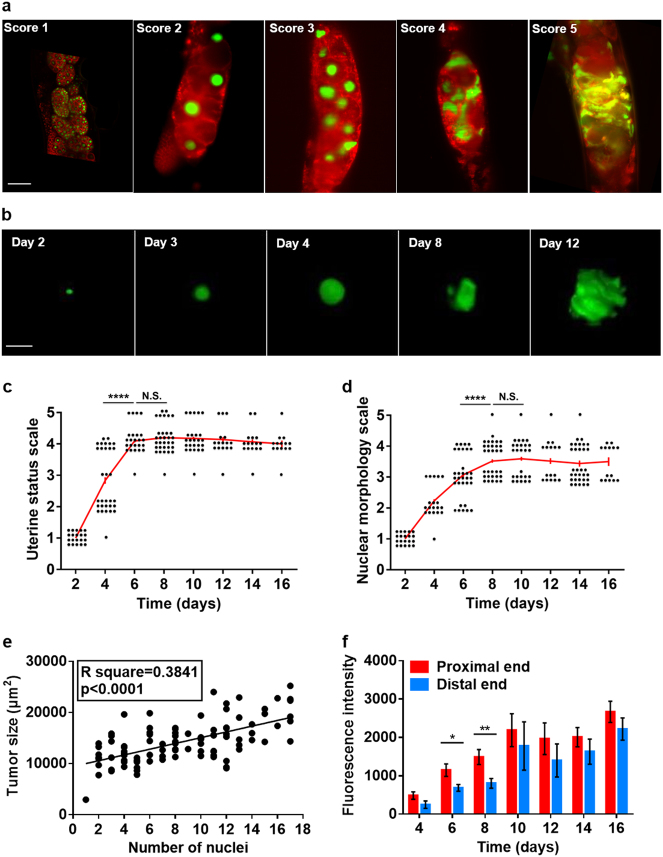
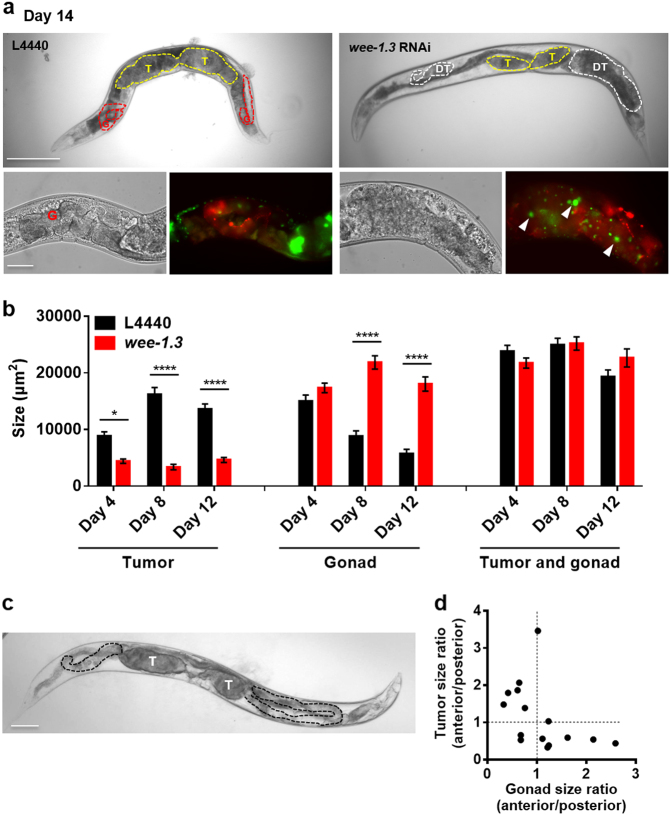
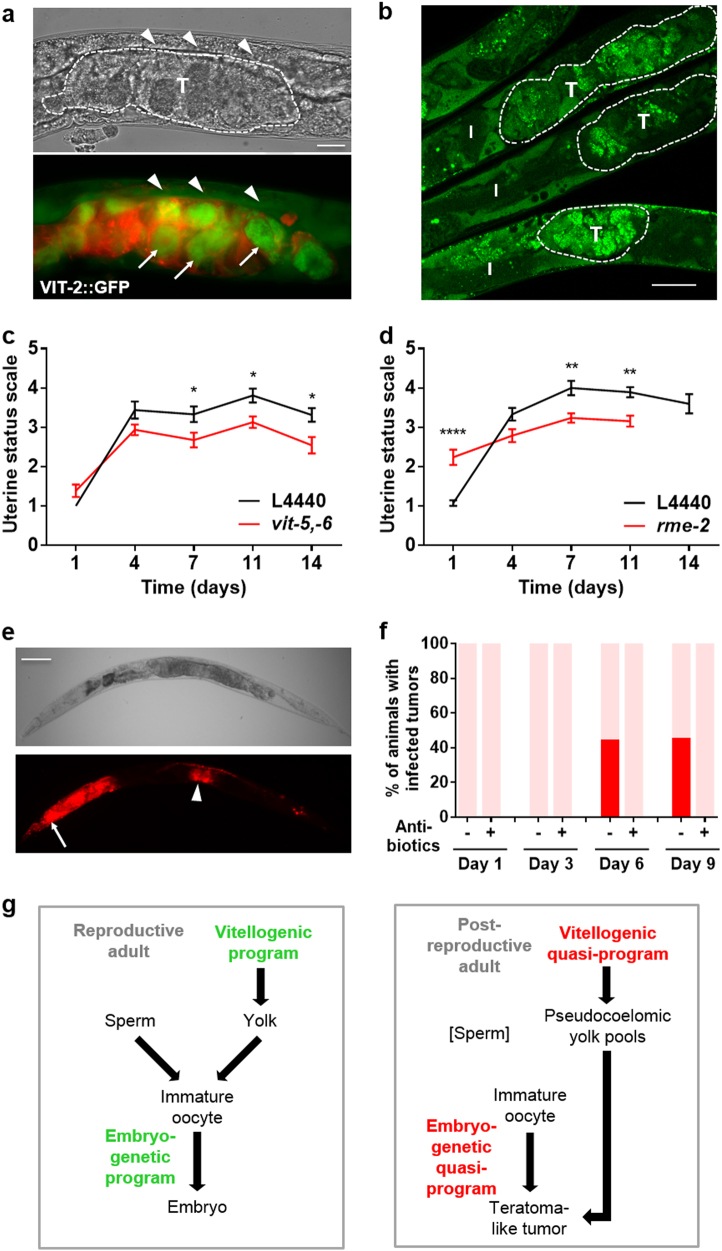
A long-standing belief is that aging (senescence) is the result of stochastic damage accumulation. Alternatively, senescent pathology may also result from late-life, wild-type gene action (i.e., antagonistic pleiotropy, as argued by Williams) leading to non-adaptive run-on of developmental programs (or quasi-programs) (as suggested more recently by Blagosklonny). In this study, we use existing and new data to show how uterine tumors, a prominent form of senescent pathology in the nematode Caenorhabditis elegans, likely result from quasi-programs. Such tumors develop from unfertilized oocytes which enter the uterus and become hypertrophic and replete with endoreduplicated chromatin masses. Tumor formation begins with ovulation of unfertilized oocytes immediately after exhaustion of sperm stocks. We show that the timing of this transition between program and quasi-program (i.e., the onset of senescence), and the onset of tumor formation, depends upon the timing of sperm depletion. We identify homology between uterine tumors and mammalian ovarian teratomas, which both develop from oocytes that fail to mature after meiosis I. In teratomas, futile activation of developmental programs leads to the formation of differentiated structures within the tumor. We report that older uterine tumors express markers of later embryogenesis, consistent with teratoma-like activation of developmental programs. We also present evidence of coupling of distal gonad atrophy to oocyte hypertrophy. This study shows how the Williams Blagosklonny model can provide a mechanistic explanation of this component of C. elegans aging. It also suggests etiological similarity between teratoma and some forms of senescent pathology, insofar as both are caused by quasi-programs.
Conflict of interest statement
The authors declare no competing interests.
Figures


References
-
- Williams GC. Pleiotropy, natural selection and the evolution of senescence. Evolution. 1957;11:398–411. doi: 10.1111/j.1558-5646.1957.tb02911.x. - DOI
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
