

Apparent calcium dependence of vesicle recruitment
- PMID: 29928766
- PMCID: PMC6166083
- DOI: 10.1113/JP275911
Apparent calcium dependence of vesicle recruitment
Abstract
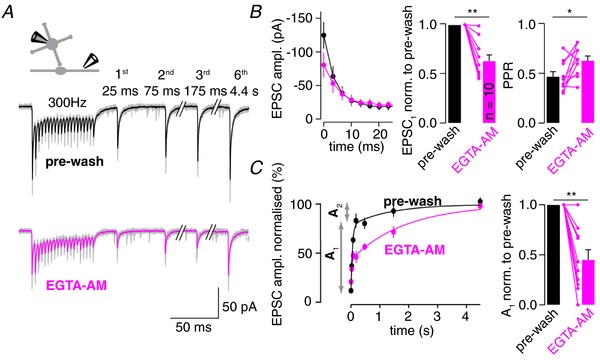
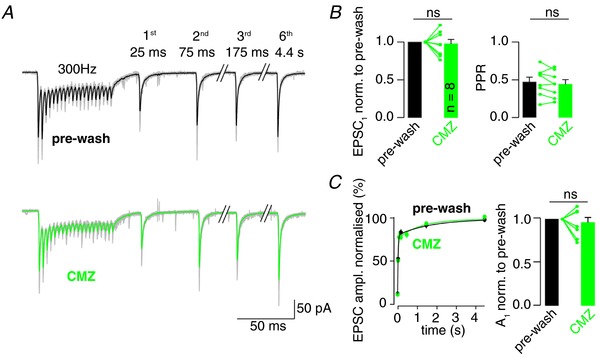
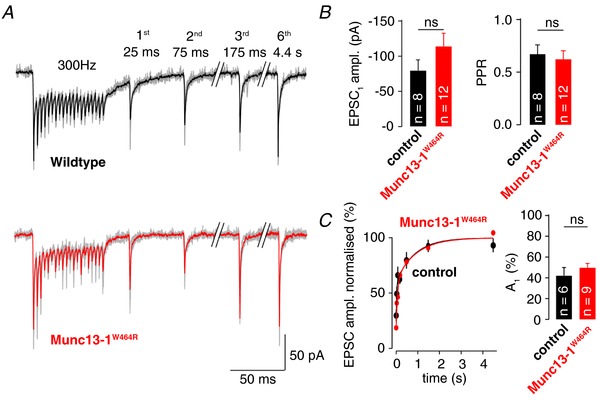
Key points: Synaptic transmission relies on the recruitment of neurotransmitter-filled vesicles to presynaptic release sites. Increased intracellular calcium buffering slows the recovery from synaptic depression, suggesting that vesicle recruitment is a calcium-dependent process. However, the molecular mechanisms of vesicle recruitment have only been investigated at some synapses. We investigate the role of calcium in vesicle recruitment at the cerebellar mossy fibre to granule cell synapse. We find that increased intracellular calcium buffering slows the recovery from depression following physiological stimulation. However, the recovery is largely resistant to perturbation of the molecular pathways previously shown to mediate calcium-dependent vesicle recruitment. Furthermore, we find two pools of vesicles with different recruitment speeds and show that models incorporating two pools of vesicles with different calcium-independent recruitment rates can explain our data. In this framework, increased calcium buffering prevents the release of intrinsically fast-recruited vesicles but does not change the vesicle recruitment rates themselves.
Abstract: During sustained synaptic transmission, recruitment of new transmitter-filled vesicles to the release site counteracts vesicle depletion and thus synaptic depression. An elevated intracellular Ca2+ concentration has been proposed to accelerate the rate of vesicle recruitment at many synapses. This conclusion is often based on the finding that increased intracellular Ca2+ buffering slows the recovery from synaptic depression. However, the molecular mechanisms of the activity-dependent acceleration of vesicle recruitment have only been analysed at some synapses. Using physiological stimulation patterns in postsynaptic recordings and step depolarizations in presynaptic bouton recordings, we investigate vesicle recruitment at cerebellar mossy fibre boutons. We show that increased intracellular Ca2+ buffering slows recovery from depression dramatically. However, pharmacological and genetic interference with calmodulin or the calmodulin-Munc13 pathway, which has been proposed to mediate Ca2+ -dependence of vesicle recruitment, barely affects vesicle recovery from depression. Furthermore, we show that cerebellar mossy fibre boutons have two pools of vesicles: rapidly fusing vesicles that recover slowly and slowly fusing vesicles that recover rapidly. Finally, models adopting such two pools of vesicles with Ca2+ -independent recruitment rates can explain the slowed recovery from depression upon increased Ca2+ buffering. Our data do not rule out the involvement of the calmodulin-Munc13 pathway during stronger stimuli or other molecular pathways mediating Ca2+ -dependent vesicle recruitment at cerebellar mossy fibre boutons. However, we show that well-established two-pool models predict an apparent Ca2+ -dependence of vesicle recruitment. Thus, previous conclusions of Ca2+ -dependent vesicle recruitment based solely on increased intracellular Ca2+ buffering should be considered with caution.
Keywords: Synapse; calcium buffering; short-term plasticity; vesicle recruitment.
© 2018 The Authors. The Journal of Physiology © 2018 The Physiological Society.
Figures



Comment in
-
Blocking slow exocytosis with slow Ca2+ buffers slows recovery from depression.J Physiol. 2018 Oct;596(19):4555-4557. doi: 10.1113/JP276673. Epub 2018 Aug 31. J Physiol. 2018. PMID: 30079526 Free PMC article. No abstract available.
References
-
- Böhme MA, Beis C, Reddy‐Alla S, Reynolds E, Mampell MM, Grasskamp AT, Lützkendorf J, Bergeron DD, Driller JH, Babikir H, Göttfert F, Robinson IM, O'Kane CJ, Hell SW, Wahl MC, Stelzl U, Loll B, Walter AM & Sigrist SJ (2016). Active zone scaffolds differentially accumulate Unc13 isoforms to tune Ca2+ channel‐vesicle coupling. Nat Neurosci 19, 1311–1320. - PubMed
-
- Borst JG & Soria van Hoeve J (2012). The calyx of Held synapse: from model synapse to auditory relay. Annu Rev Physiol 74, 199–224. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous