

Trim33 mediates the proinflammatory function of Th17 cells
- PMID: 29930104
- PMCID: PMC6028517
- DOI: 10.1084/jem.20170779
Trim33 mediates the proinflammatory function of Th17 cells
Abstract
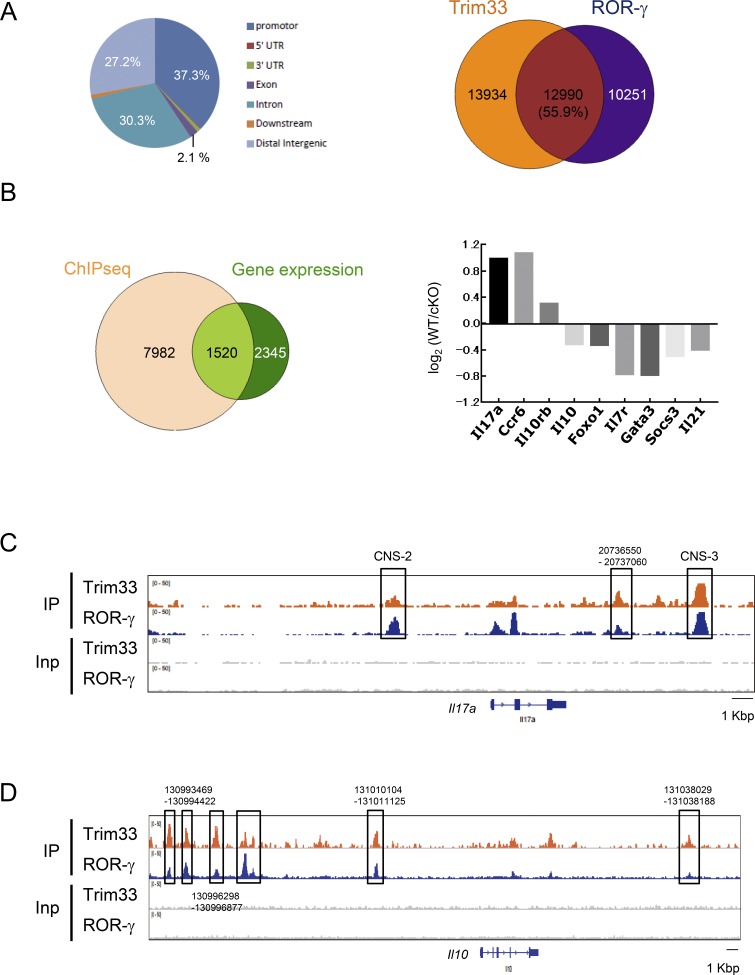
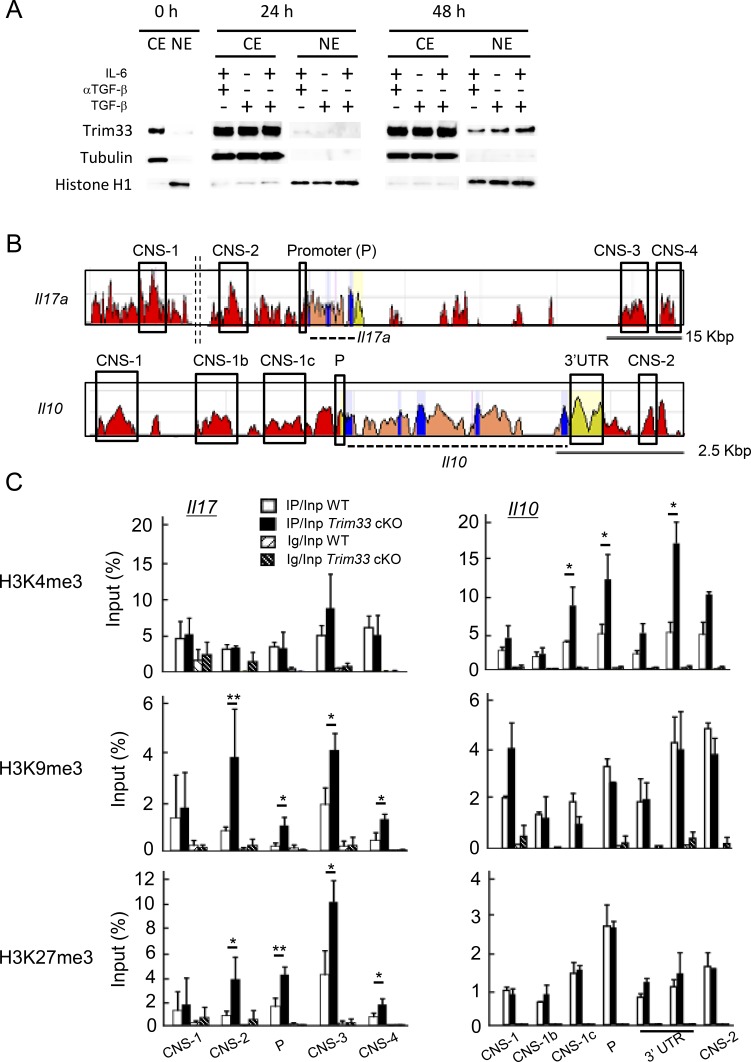
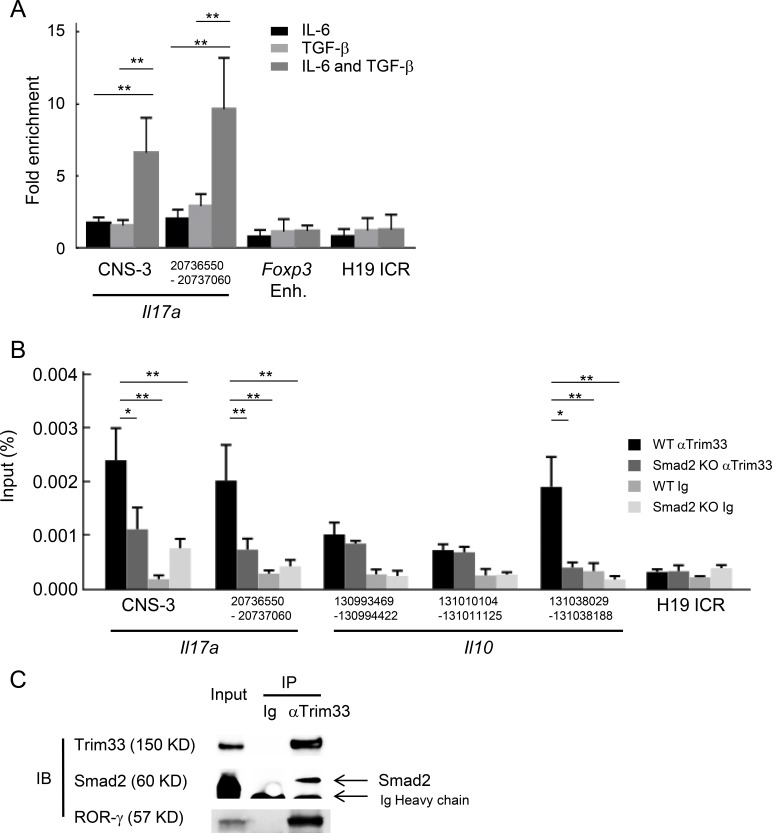
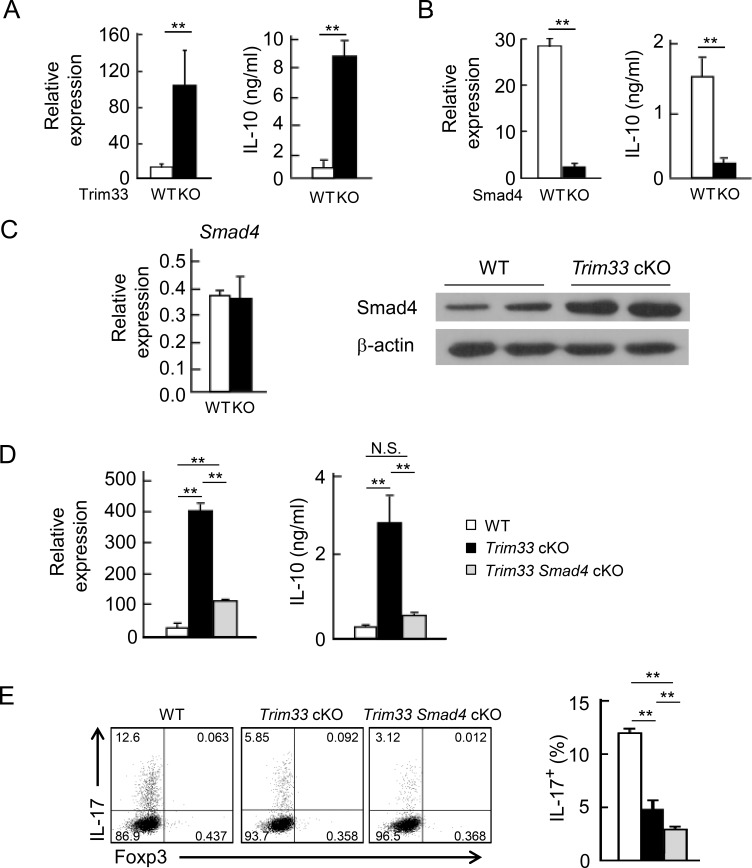
Transforming growth factor-β (TGF-β) regulates reciprocal regulatory T cell (T reg) and T helper 17 (Th17) differentiation, the underlying mechanism of which is still not understood. Here, we report that tripartite motif-containing 33 (Trim33), a modulator of TGF-β signaling that associates with Smad2, regulates the proinflammatory function of Th17 cells. Trim33 deficiency in T cells ameliorated an autoimmune disease in vivo. Trim33 was required for induction in vitro of Th17, but not T reg cells. Moreover, Smad4 and Trim33 play contrasting roles in the regulation of IL-10 expression; loss of Trim33 enhanced IL-10 production. Furthermore, Trim33 was recruited to the Il17a and Il10 gene loci, dependent on Smad2, and mediated their chromatin remodeling during Th17 differentiation. Trim33 thus promotes the proinflammatory function of Th17 cells by inducing IL-17 and suppressing IL-10 expression.
© 2018 Tanaka et al.
Figures
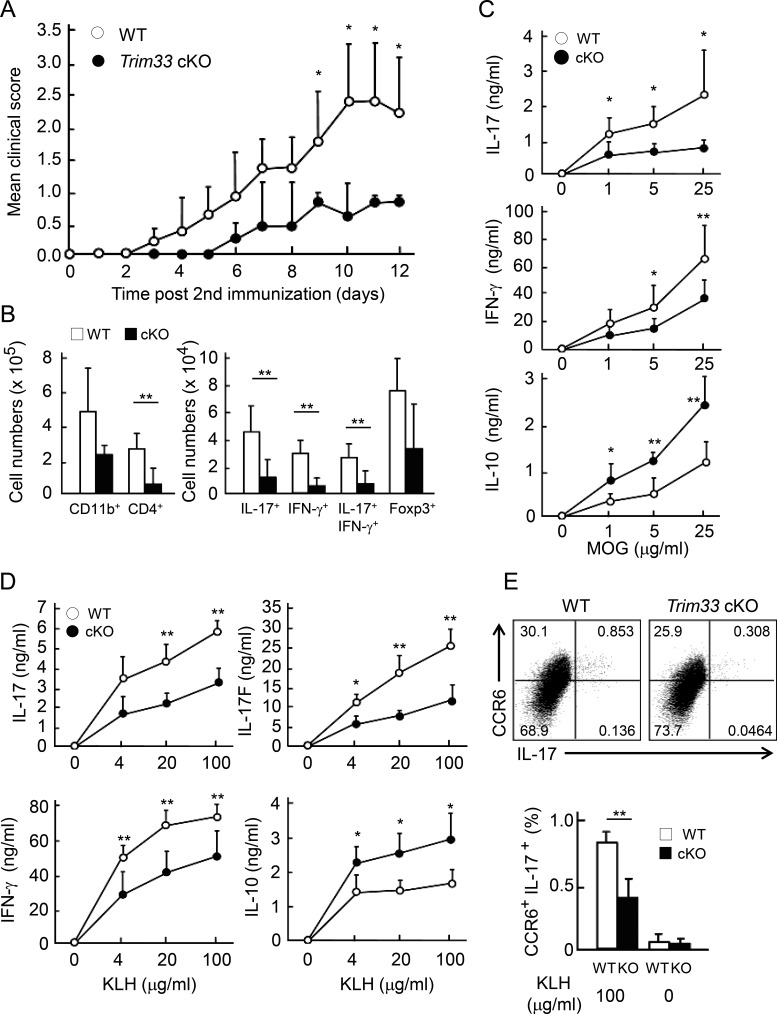
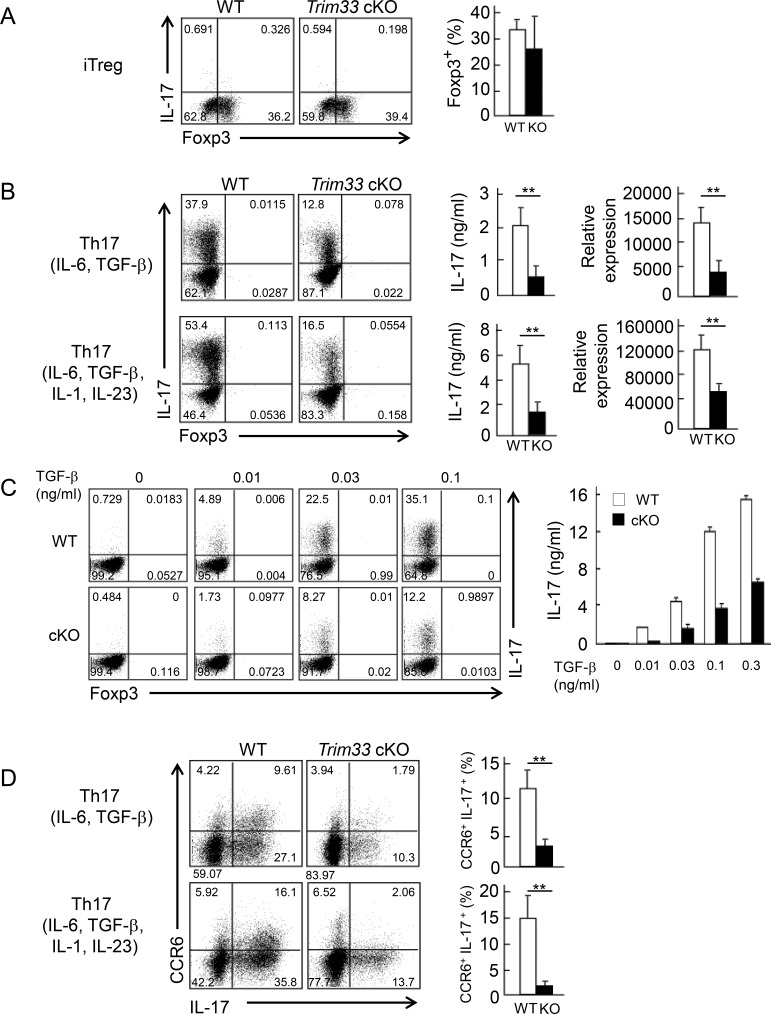
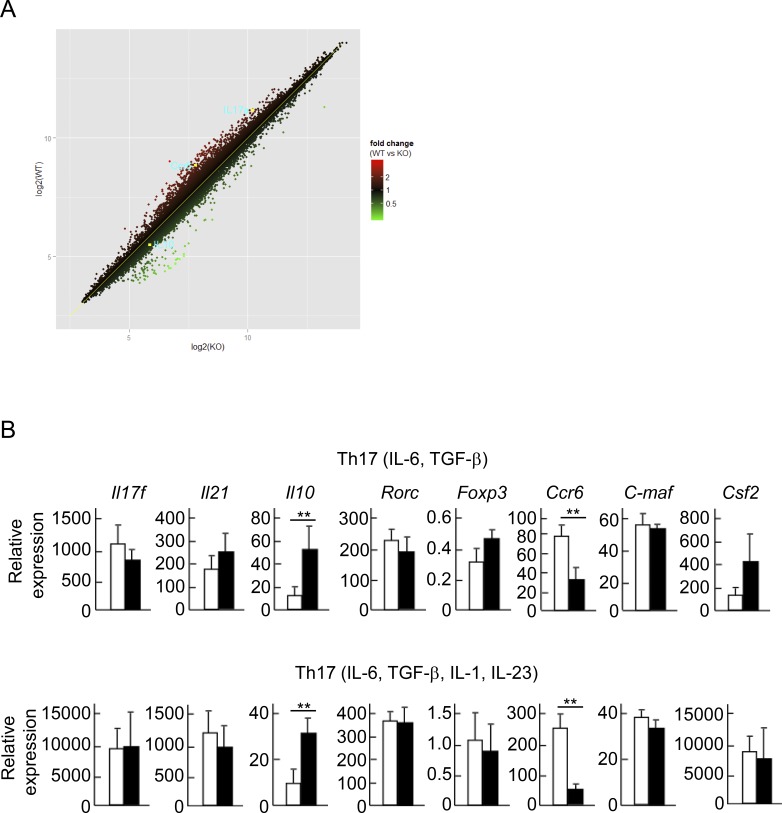
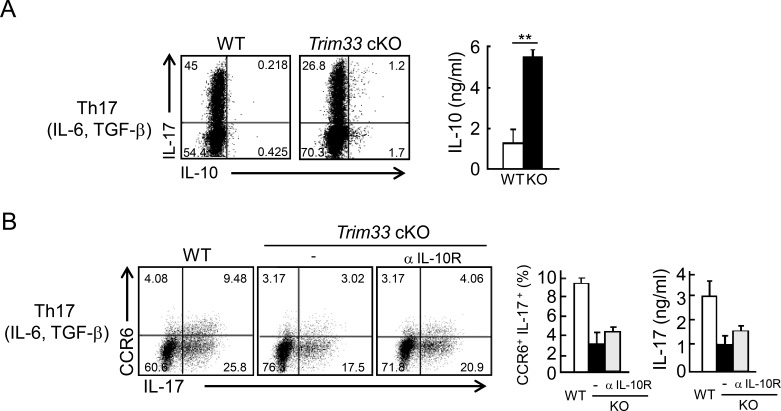
Comment in
-
TRIMming TGF-β signals in Th17 cells.J Exp Med. 2018 Jul 2;215(7):1775-1776. doi: 10.1084/jem.20180986. Epub 2018 Jun 26. J Exp Med. 2018. PMID: 29945999 Free PMC article.
References
-
- Bettelli E., Das M.P., Howard E.D., Weiner H.L., Sobel R.A., and Kuchroo V.K.. 1998. IL-10 is critical in the regulation of autoimmune encephalomyelitis as demonstrated by studies of IL-10- and IL-4-deficient and transgenic mice. J. Immunol. 161:3299–3306. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
