

Cytosolic Hsp70 and Hsp40 chaperones enable the biogenesis of mitochondrial β-barrel proteins
- PMID: 29930205
- PMCID: PMC6122992
- DOI: 10.1083/jcb.201712029
Cytosolic Hsp70 and Hsp40 chaperones enable the biogenesis of mitochondrial β-barrel proteins
Abstract
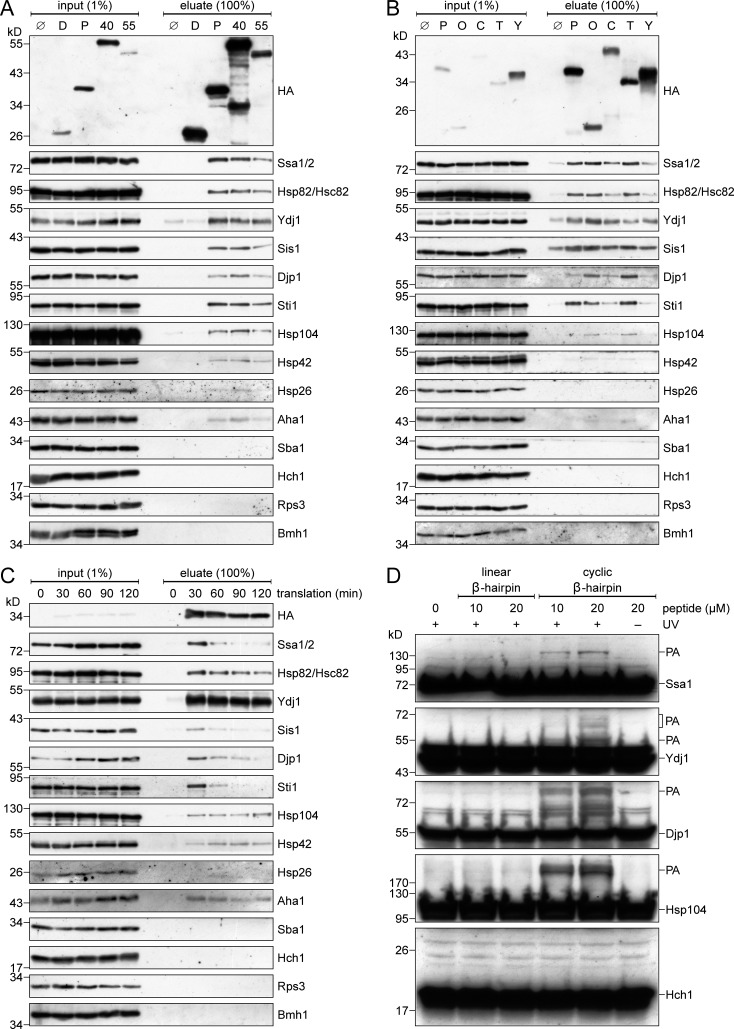
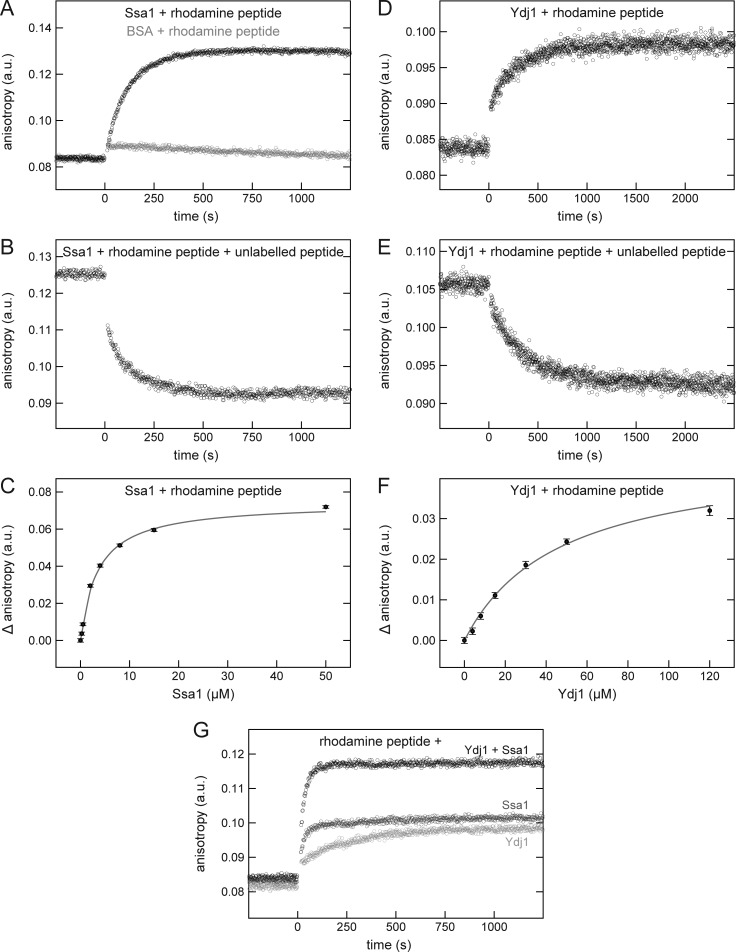
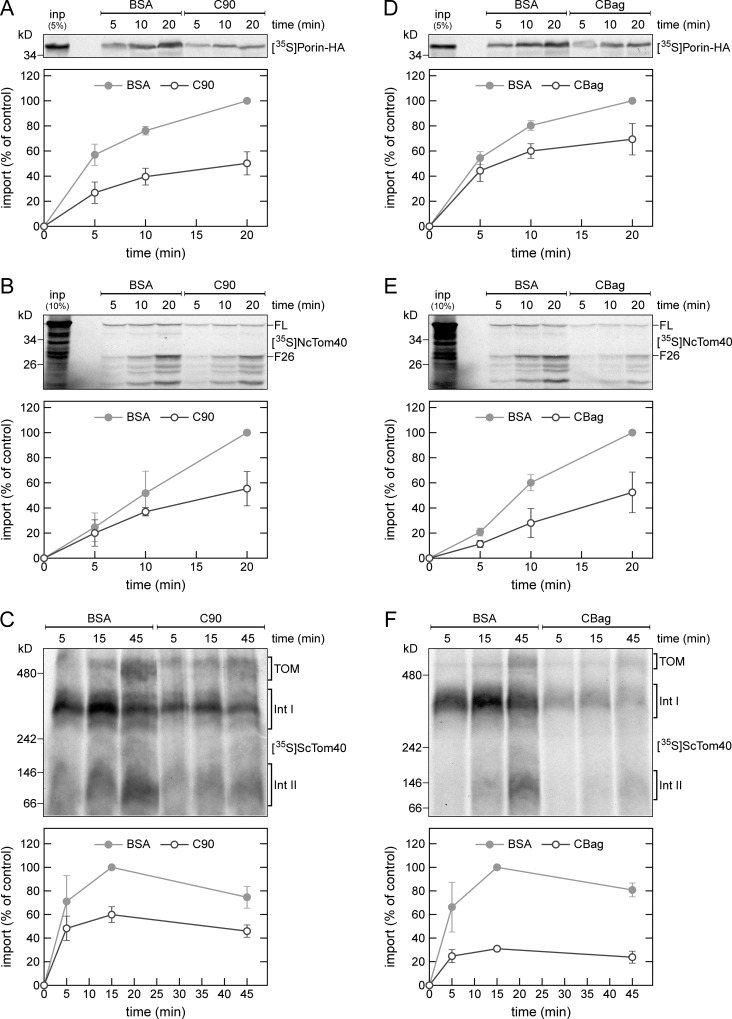
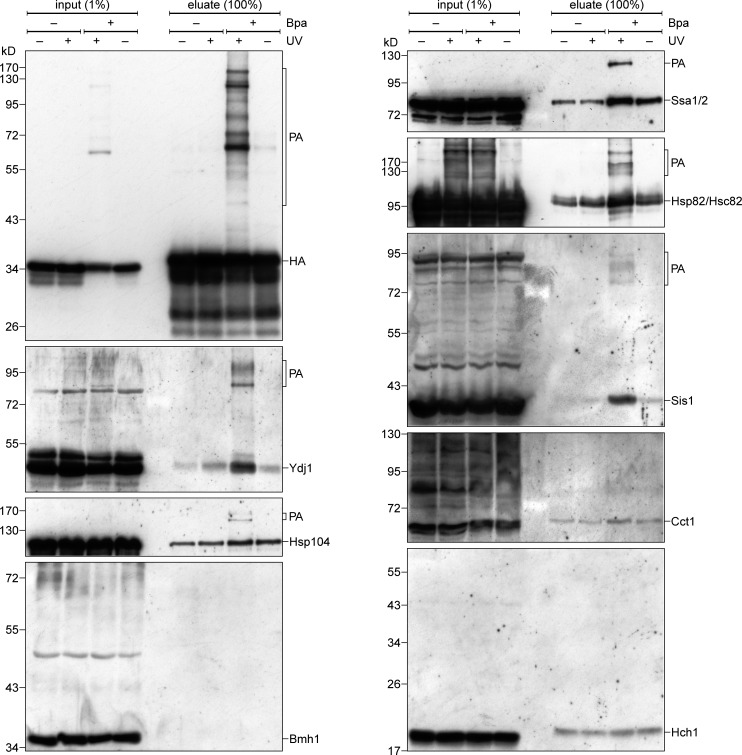
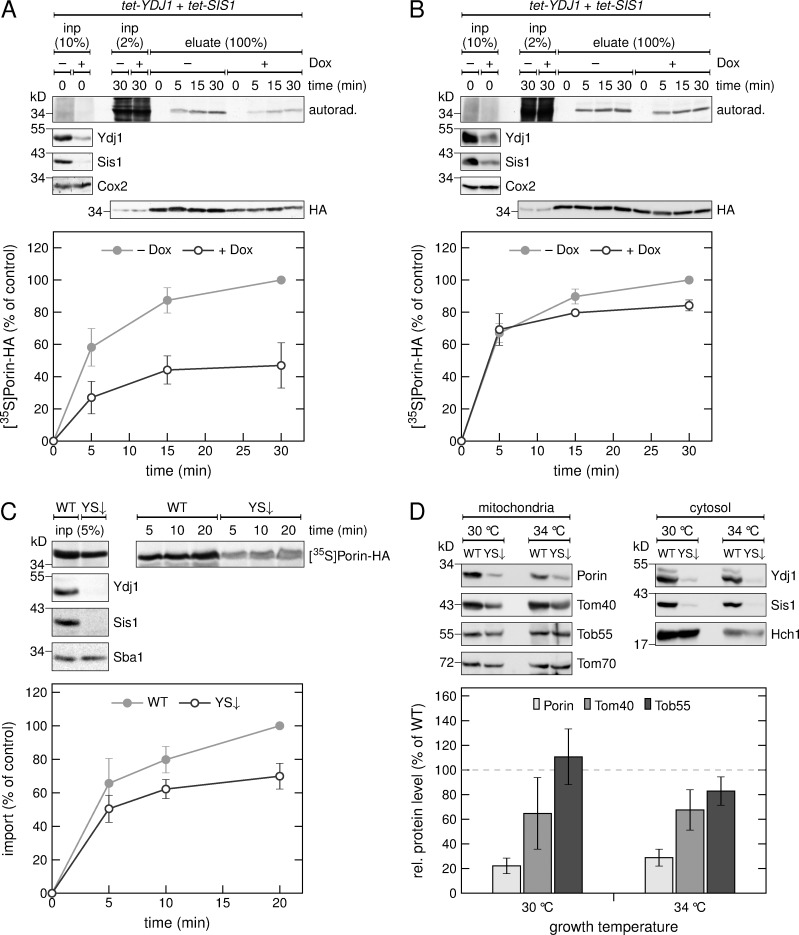
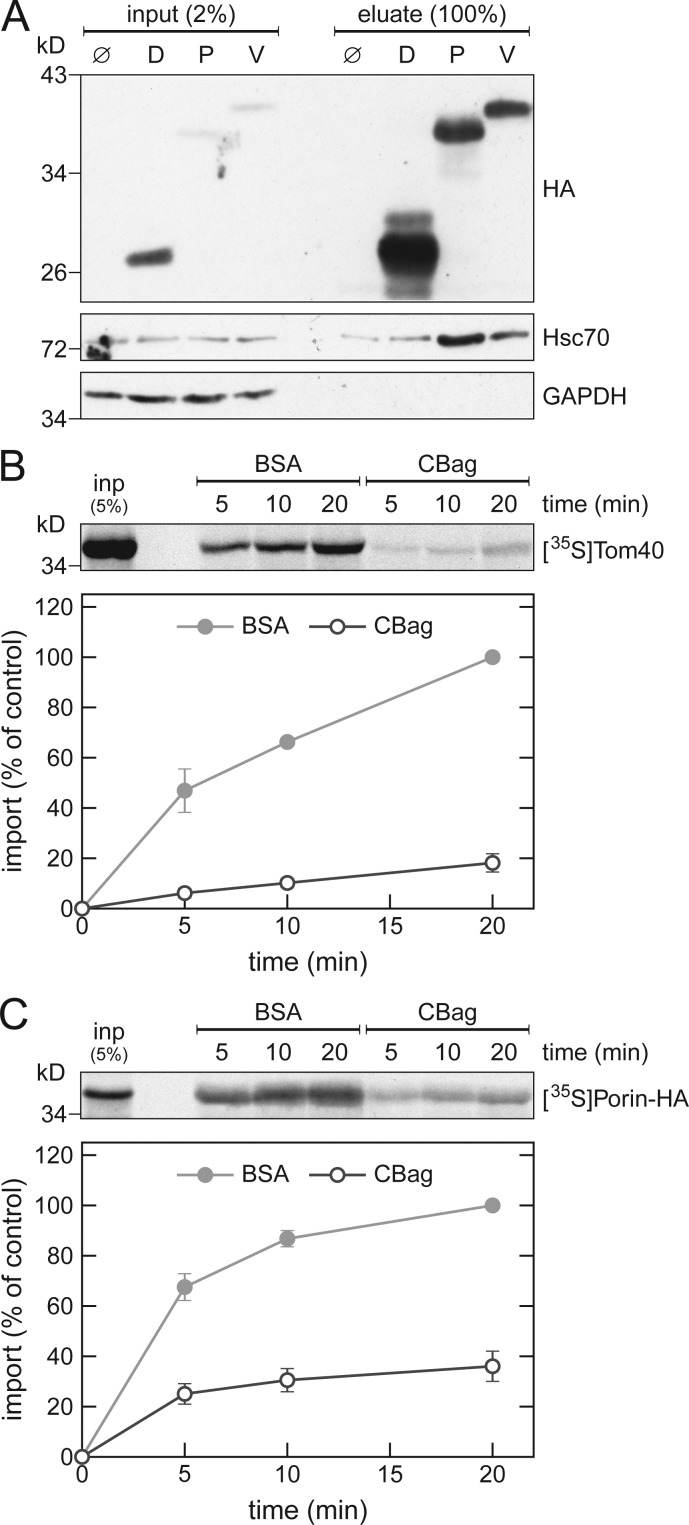
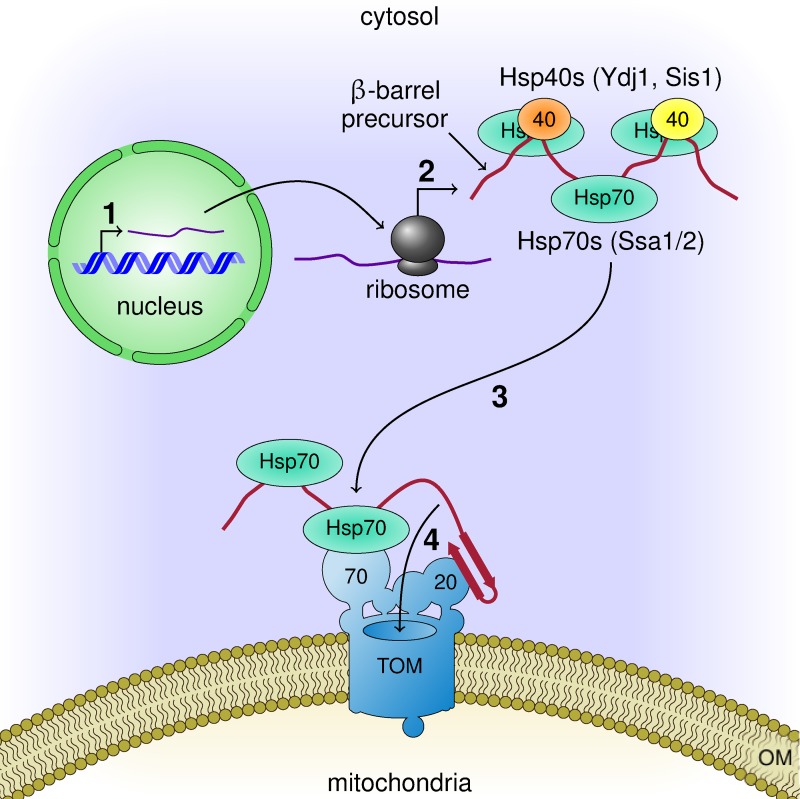
Mitochondrial β-barrel proteins are encoded in the nucleus, translated by cytosolic ribosomes, and then imported into the organelle. Recently, a detailed understanding of the intramitochondrial import pathway of β-barrel proteins was obtained. In contrast, it is still completely unclear how newly synthesized β-barrel proteins reach the mitochondrial surface in an import-competent conformation. In this study, we show that cytosolic Hsp70 chaperones and their Hsp40 cochaperones Ydj1 and Sis1 interact with newly synthesized β-barrel proteins. These interactions are highly relevant for proper biogenesis, as inhibiting the activity of the cytosolic Hsp70, preventing its docking to the mitochondrial receptor Tom70, or depleting both Ydj1 and Sis1 resulted in a significant reduction in the import of such substrates into mitochondria. Further experiments demonstrate that the interactions between β-barrel proteins and Hsp70 chaperones and their importance are conserved also in mammalian cells. Collectively, this study outlines a novel mechanism in the early events of the biogenesis of mitochondrial outer membrane β-barrel proteins.
© 2018 Jores et al.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
