

The evolution of molluscs
- PMID: 29931833
- PMCID: PMC6378612
- DOI: 10.1111/brv.12439
The evolution of molluscs
Abstract
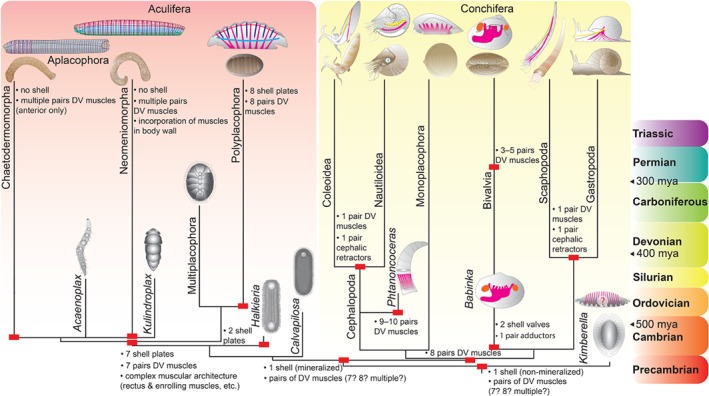
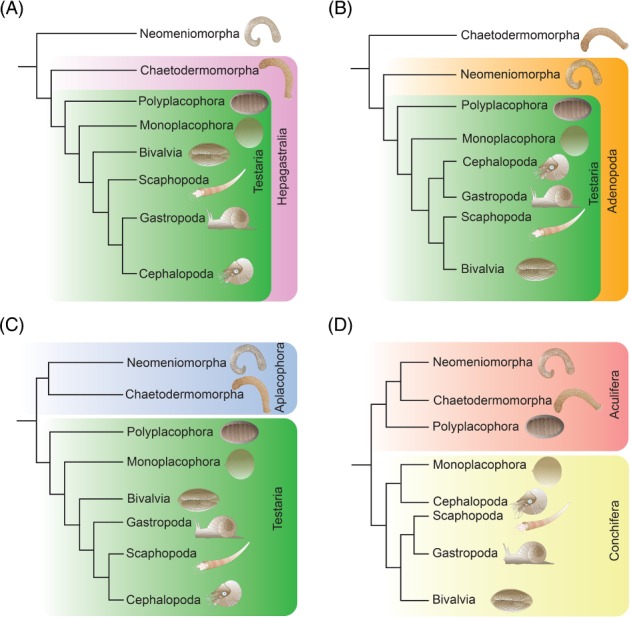
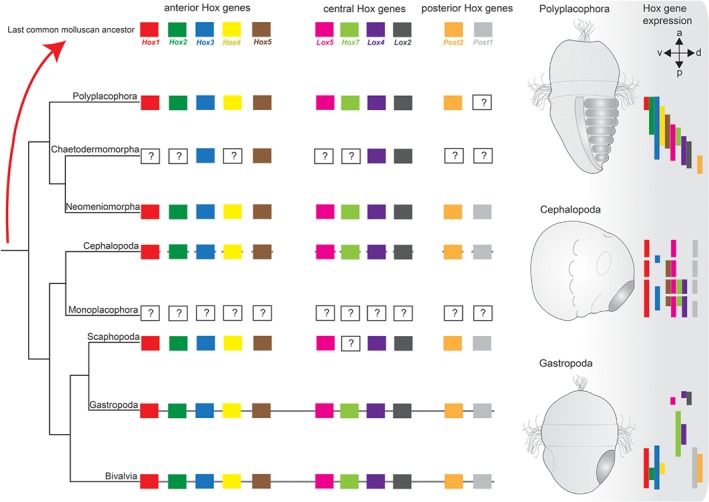
Molluscs are extremely diverse invertebrate animals with a rich fossil record, highly divergent life cycles, and considerable economical and ecological importance. Key representatives include worm-like aplacophorans, armoured groups (e.g. polyplacophorans, gastropods, bivalves) and the highly complex cephalopods. Molluscan origins and evolution of their different phenotypes have largely remained unresolved, but significant progress has been made over recent years. Phylogenomic studies revealed a dichotomy of the phylum, resulting in Aculifera (shell-less aplacophorans and multi-shelled polyplacophorans) and Conchifera (all other, primarily uni-shelled groups). This challenged traditional hypotheses that proposed that molluscs gradually evolved complex phenotypes from simple, worm-like animals, a view that is corroborated by developmental studies that showed that aplacophorans are secondarily simplified. Gene expression data indicate that key regulators involved in anterior-posterior patterning (the homeobox-containing Hox genes) lost this function and were co-opted into the evolution of taxon-specific novelties in conchiferans. While the bone morphogenetic protein (BMP)/decapentaplegic (Dpp) signalling pathway, that mediates dorso-ventral axis formation, and molecular components that establish chirality appear to be more conserved between molluscs and other metazoans, variations from the common scheme occur within molluscan sublineages. The deviation of various molluscs from developmental pathways that otherwise appear widely conserved among metazoans provides novel hypotheses on molluscan evolution that can be tested with genome editing tools such as the CRISPR/Cas9 (clustered regularly interspaced short palindromic repeats/clustered regularly interspaced short palindromic repeats-associated protein9) system.
Keywords: BMP; Bilateria; Cambrian; EvoDevo; Hox; Lophotrochozoa; Mollusca; body axis; development; morphology.
© 2018 The Authors. Biological Reviews published by John Wiley ' Sons Ltd on behalf of Cambridge Philosophical Society.
Figures
References
-
- Adema, C. M. , Hillier, L. W. , Jones, C. S. , Loker, E. S. , Knight, M. , Minx, P. , Oliveira, G. , Raghavan, N. , Shedlock, A. , Rodrigues do Amaral, L. , Arican‐Goktas, H. D. , Assis, J. G. , Hideo Baba, E. , Baron, O. L. , Bayne, C. J. , et al. (2017). Whole genome analysis of a schistosomiasis‐transmitting freshwater snail. Nature Communications 8, 15451. - PMC - PubMed
-
- Anderson, P. D. & Bokor, G. (2012). Conotoxins: potential weapons from the sea. Journal of Bioterrorism and Biodefence 3, 120.
-
- Anderson, G. J. & Darshan, D. (2008). Small‐molecule dissection of BMP signaling. Nature Chemical Biology 4, 15–16. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
