

Plastidial NAD-Dependent Malate Dehydrogenase: A Moonlighting Protein Involved in Early Chloroplast Development through Its Interaction with an FtsH12-FtsHi Protease Complex
- PMID: 29934433
- PMCID: PMC6139691
- DOI: 10.1105/tpc.18.00121
Plastidial NAD-Dependent Malate Dehydrogenase: A Moonlighting Protein Involved in Early Chloroplast Development through Its Interaction with an FtsH12-FtsHi Protease Complex
Abstract
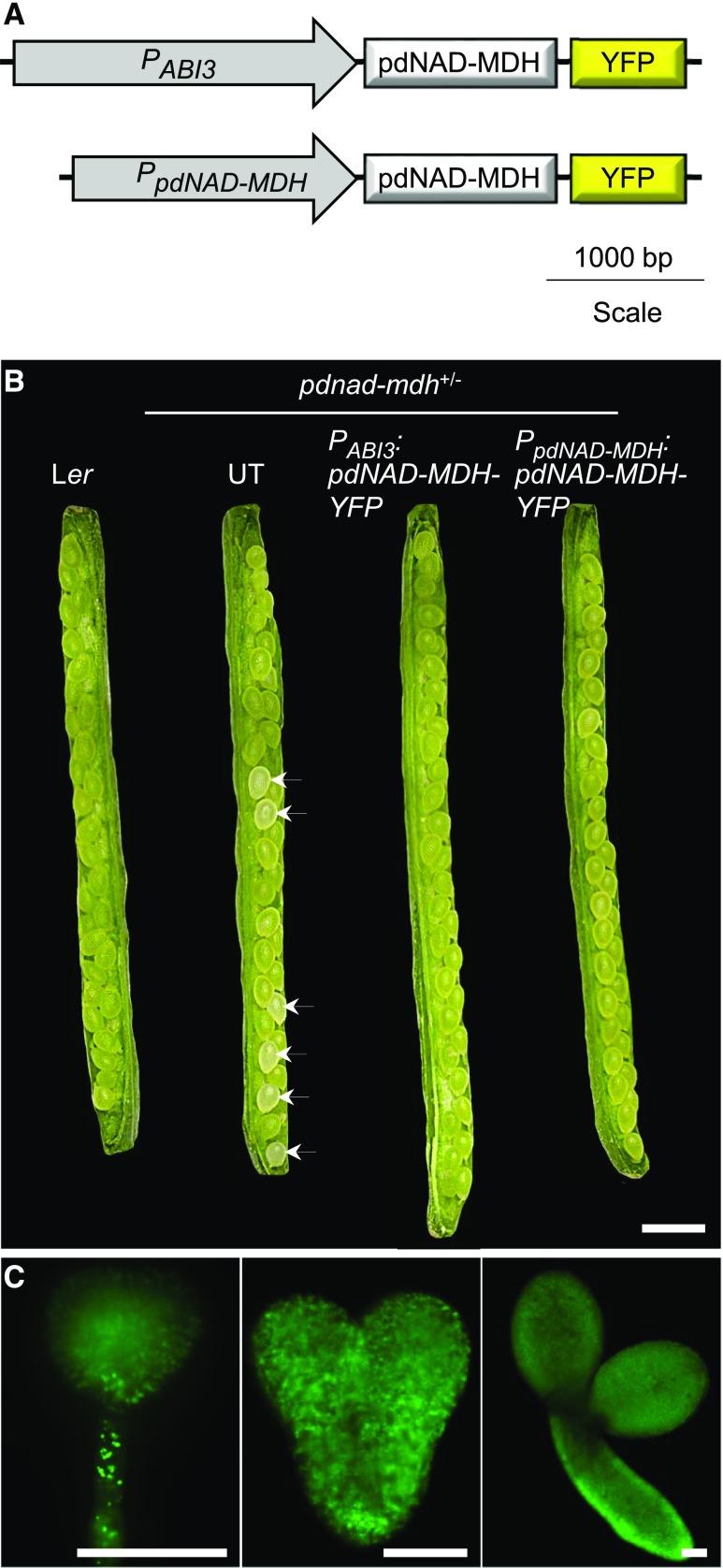
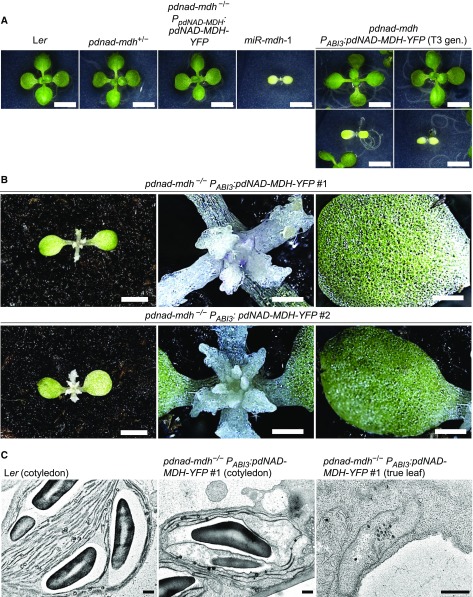
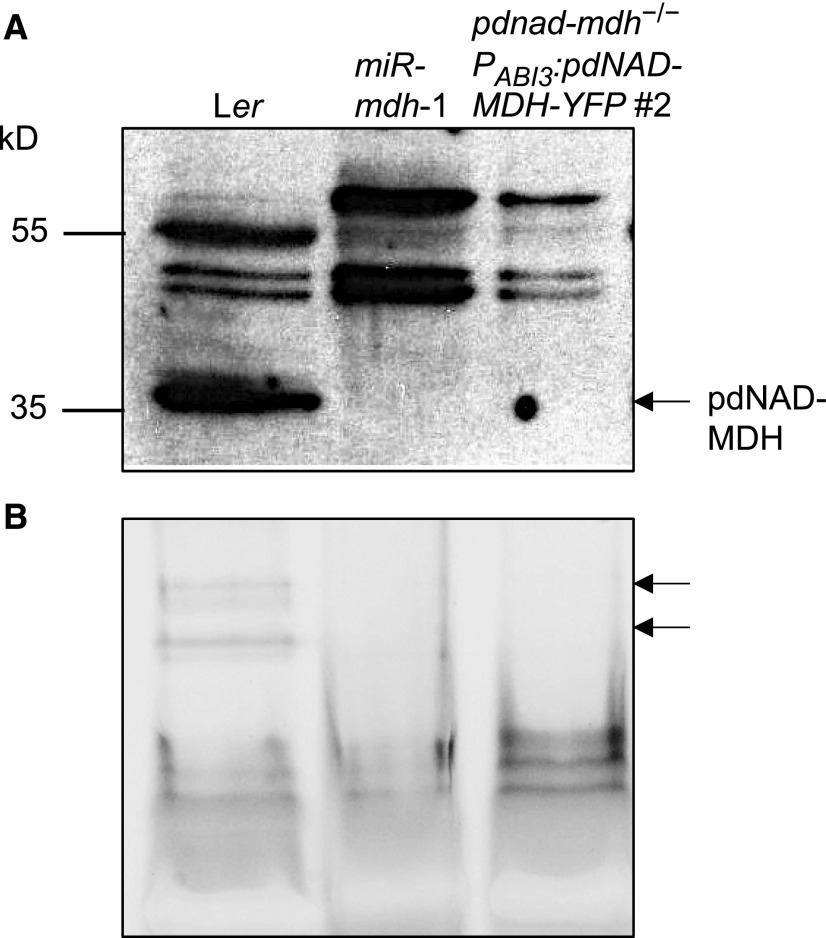
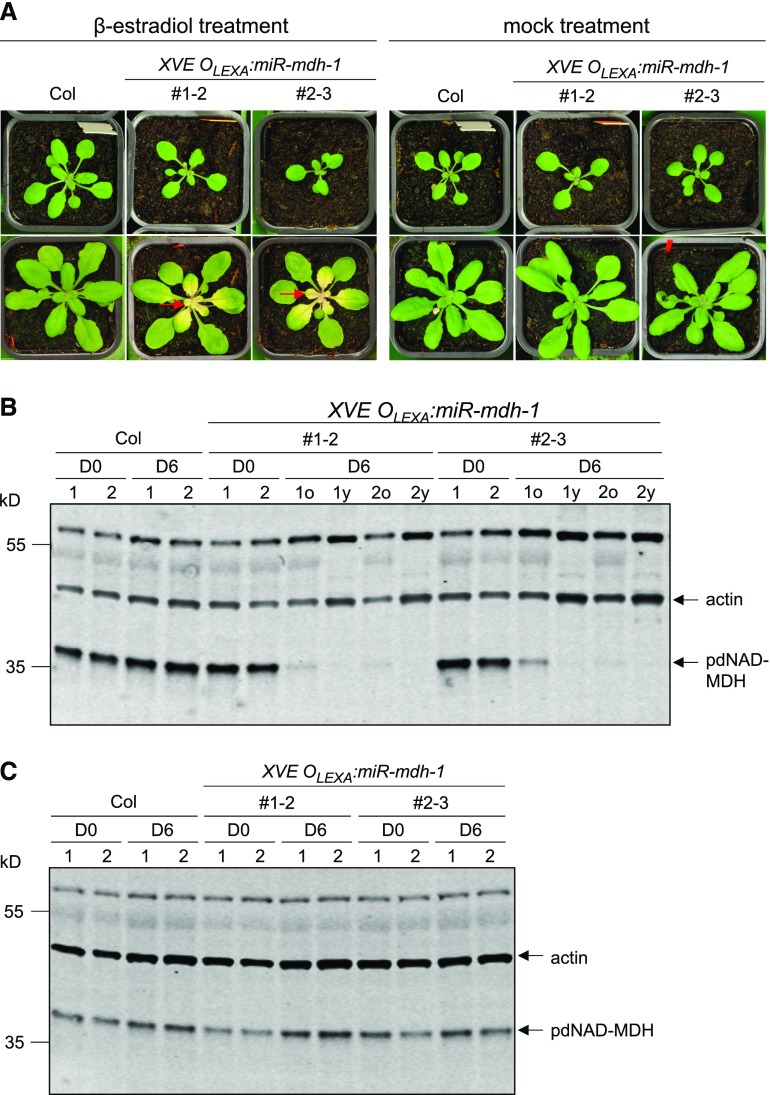
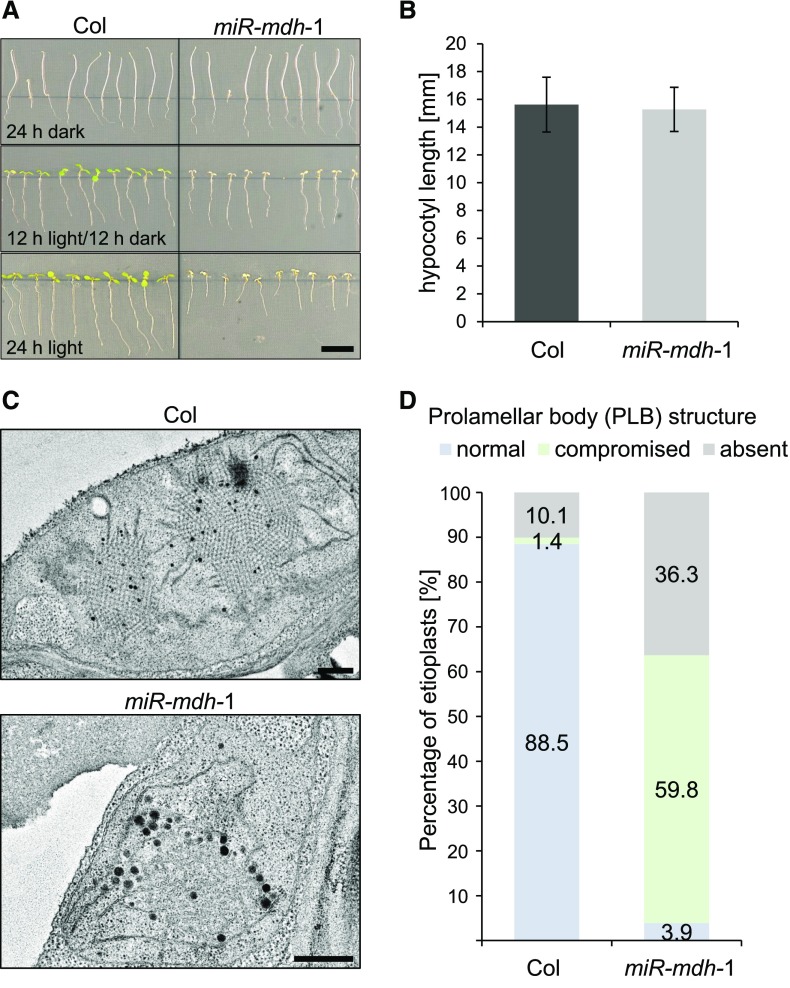
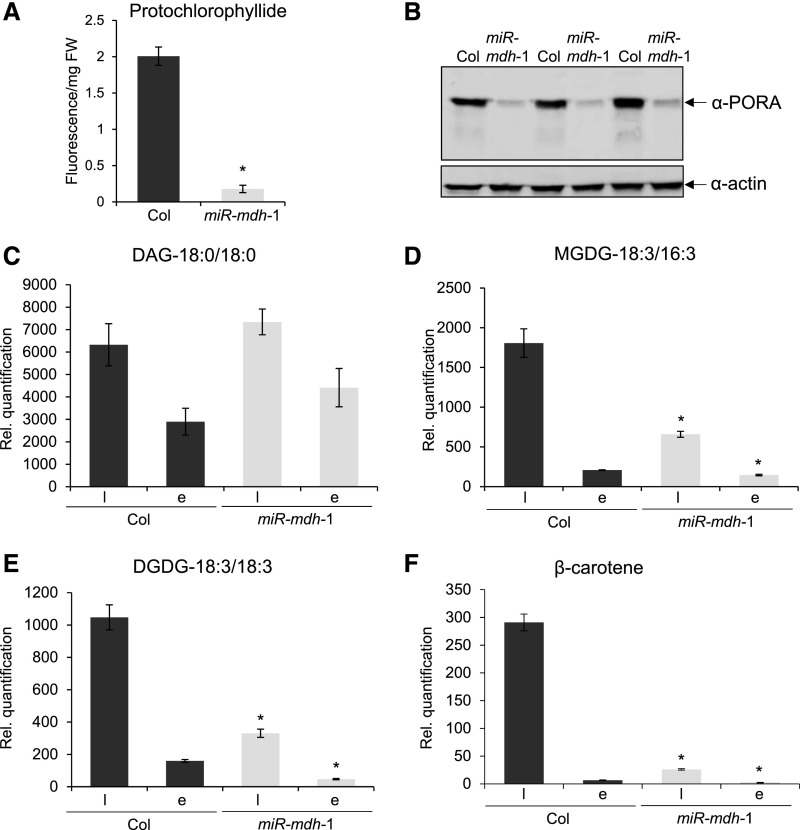
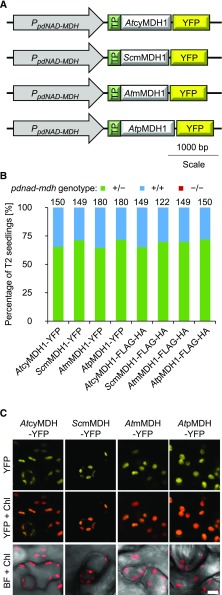
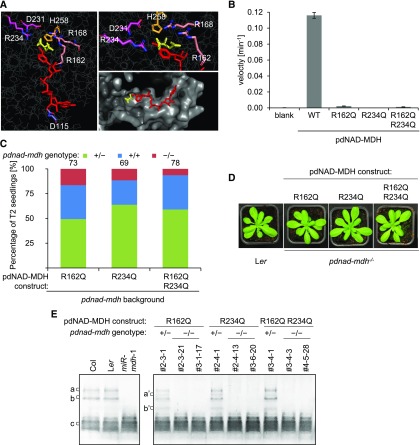
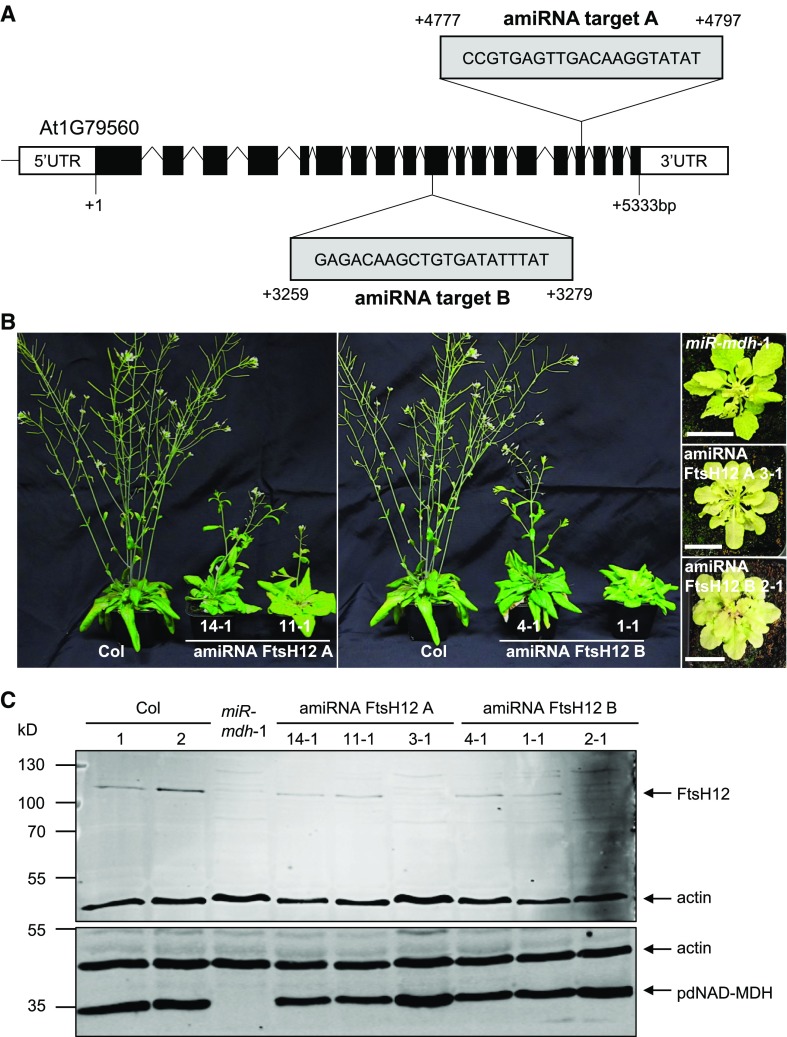
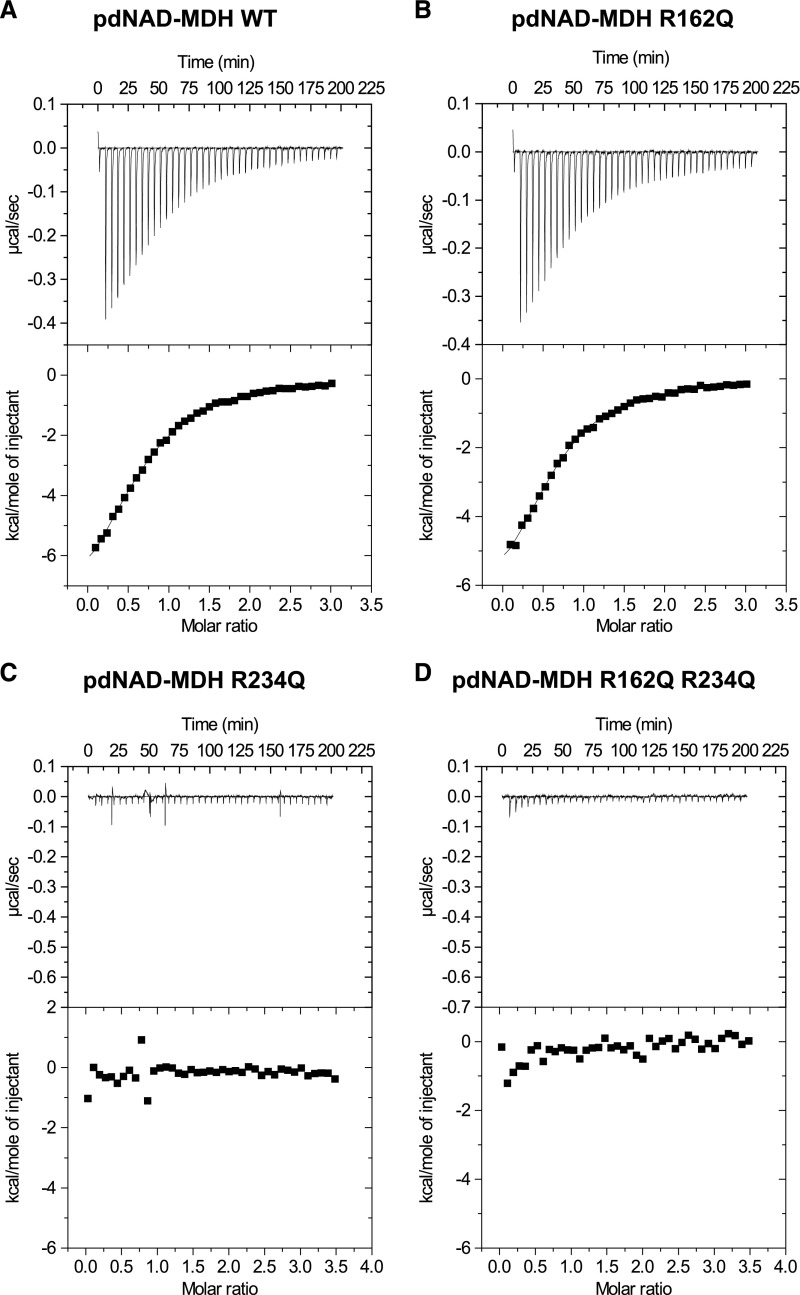
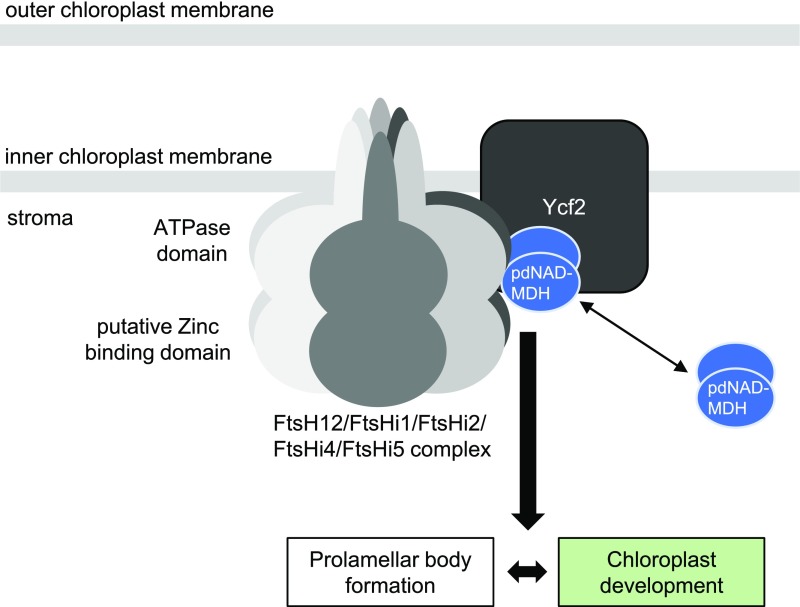
Malate dehydrogenases (MDHs) convert malate to oxaloacetate using NAD(H) or NADP(H) as a cofactor. Arabidopsis thaliana mutants lacking plastidial NAD-dependent MDH (pdnad-mdh) are embryo-lethal, and constitutive silencing (miR-mdh-1) causes a pale, dwarfed phenotype. The reason for these severe phenotypes is unknown. Here, we rescued the embryo lethality of pdnad-mdh via embryo-specific expression of pdNAD-MDH. Rescued seedlings developed white leaves with aberrant chloroplasts and failed to reproduce. Inducible silencing of pdNAD-MDH at the rosette stage also resulted in white newly emerging leaves. These data suggest that pdNAD-MDH is important for early plastid development, which is consistent with the reductions in major plastidial galactolipid, carotenoid, and protochlorophyllide levels in miR-mdh-1 seedlings. Surprisingly, the targeting of other NAD-dependent MDH isoforms to the plastid did not complement the embryo lethality of pdnad-mdh, while expression of enzymatically inactive pdNAD-MDH did. These complemented plants grew indistinguishably from the wild type. Both active and inactive forms of pdNAD-MDH interact with a heteromeric AAA-ATPase complex at the inner membrane of the chloroplast envelope. Silencing the expression of FtsH12, a key member of this complex, resulted in a phenotype that strongly resembles miR-mdh-1. We propose that pdNAD-MDH is essential for chloroplast development due to its moonlighting role in stabilizing FtsH12, distinct from its enzymatic function.
© 2018 American Society of Plant Biologists. All rights reserved.
Figures

Comment in
-
Moonlighting NAD+ Malate Dehydrogenase Is Essential for Chloroplast Biogenesis.Plant Cell. 2018 Aug;30(8):1663-1664. doi: 10.1105/tpc.18.00474. Epub 2018 Jun 22. Plant Cell. 2018. PMID: 29934434 Free PMC article. No abstract available.
References
-
- Bahl J., Francke B., Monéger R. (1976). Lipid composition of envelopes, prolamellar bodies and other plastid membranes in etiolated, green and greening wheat leaves. Planta 129: 193–201. - PubMed
-
- Berkemeyer M., Scheibe R., Ocheretina O. (1998). A novel, non-redox-regulated NAD-dependent malate dehydrogenase from chloroplasts of Arabidopsis thaliana L. J. Biol. Chem. 273: 27927–27933. - PubMed
-
- Birktoft J.J., Banaszak L.J. (1983). The presence of a histidine-aspartic acid pair in the active site of 2-hydroxyacid dehydrogenases. X-ray refinement of cytoplasmic malate dehydrogenase. J. Biol. Chem. 258: 472–482. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
