

Pyruvate dehydrogenase complex plays a central role in brown adipocyte energy expenditure and fuel utilization during short-term beta-adrenergic activation
- PMID: 29934543
- PMCID: PMC6015083
- DOI: 10.1038/s41598-018-27875-3
Pyruvate dehydrogenase complex plays a central role in brown adipocyte energy expenditure and fuel utilization during short-term beta-adrenergic activation
Abstract
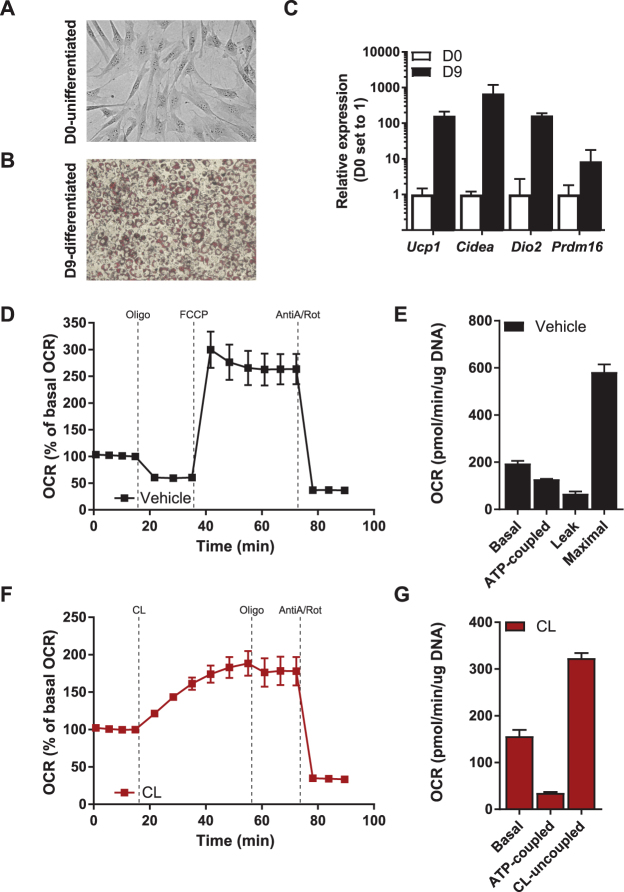
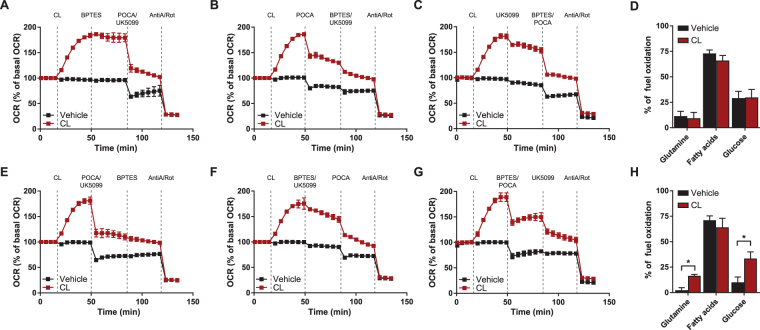
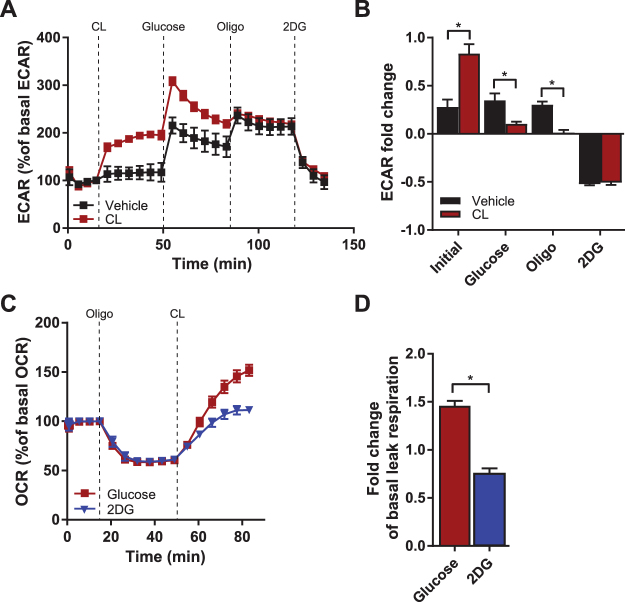
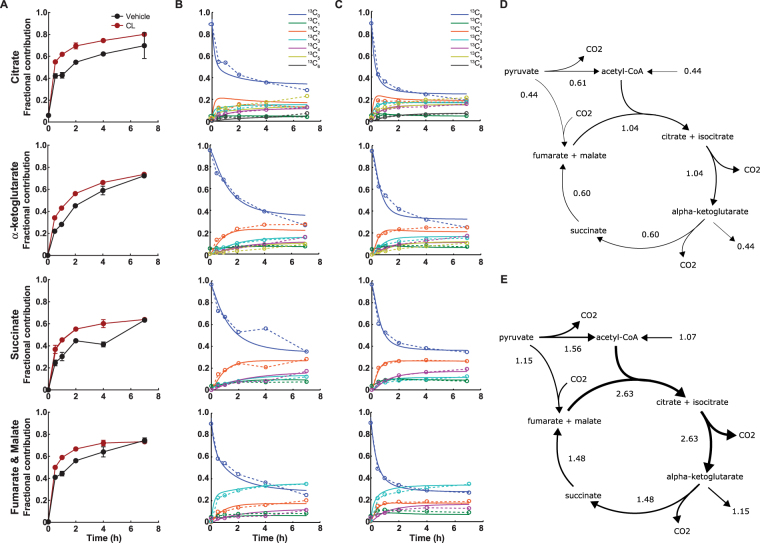
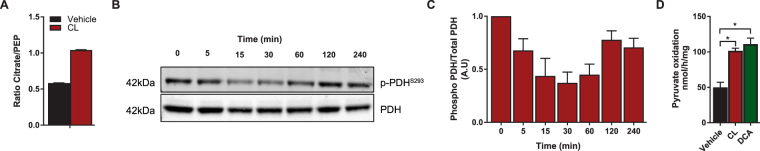
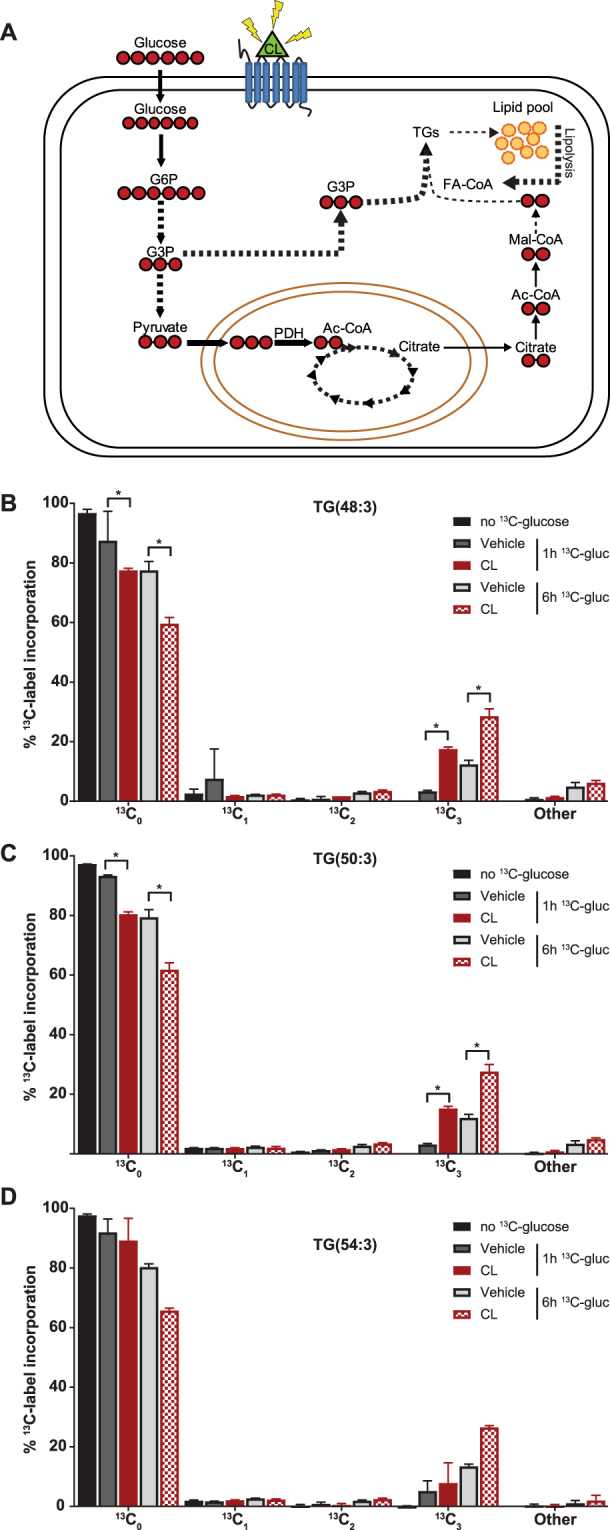
Activation of brown adipose tissue (BAT) contributes to total body energy expenditure through energy dissipation as heat. Activated BAT increases the clearance of lipids and glucose from the circulation, but how BAT accommodates large influx of multiple substrates is not well defined. The purpose of this work was to assess the metabolic fluxes in brown adipocytes during β3-adrenergic receptor (β3-AR) activation.T37i murine preadipocytes were differentiated into brown adipocytes and we used Seahorse respirometry employing a set of specific substrate inhibitors in the presence or absence of β3-AR agonist CL316,243. The main substrate used by these brown adipocytes were fatty acids, which were oxidized equally during activation as well as during resting condition. [U-13C]-glucose tracer-based metabolomics revealed that the flux through the TCA cycle was enhanced and regulated by pyruvate dehydrogenase (PDH) activity. Based on 13C-tracer incorporation in lipids, it appeared that most glucose was oxidized via TCA cycle activity, while some was utilized for glycerol-3-phosphate synthesis to replenish the triglyceride pool. Collectively, we show that while fatty acids are the main substrates for oxidation, glucose is also oxidized to meet the increased energy demand during short term β3-AR activation. PDH plays an important role in directing glucose carbons towards oxidation.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
