

Mechanical transduction of cytoplasmic-to-transmembrane-domain movements in a hyperpolarization-activated cyclic nucleotide-gated cation channel
- PMID: 29936413
- PMCID: PMC6102142
- DOI: 10.1074/jbc.RA118.002139
Mechanical transduction of cytoplasmic-to-transmembrane-domain movements in a hyperpolarization-activated cyclic nucleotide-gated cation channel
Abstract
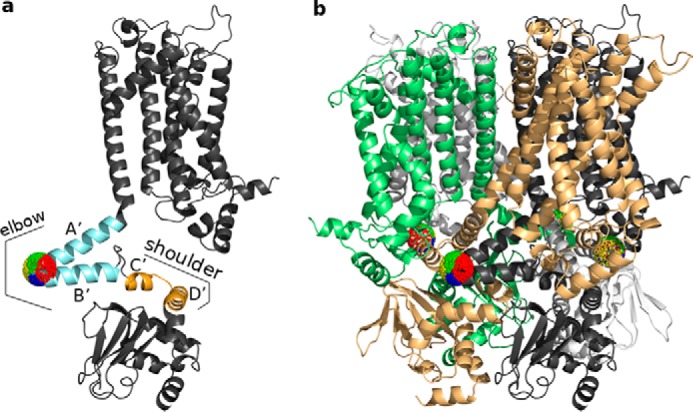
Hyperpolarization-activated cyclic nucleotide-gated cation (HCN) channels play a critical role in the control of pacemaking in the heart and repetitive firing in neurons. In HCN channels, the intracellular cyclic nucleotide-binding domain (CNBD) is connected to the transmembrane portion of the channel (TMPC) through a helical domain, the C-linker. Although this domain is critical for mechanical signal transduction, the conformational dynamics in the C-linker that transmit the nucleotide-binding signal to the HCN channel pore are unknown. Here, we use linear response theory to analyze conformational changes in the C-linker of the human HCN1 protein, which couple cAMP binding in the CNBD with gating in the TMPC. By applying a force to the tip of the so-called "elbow" of the C-linker, the coarse-grained calculations recapitulate the same conformational changes triggered by cAMP binding in experimental studies. Furthermore, in our simulations, a displacement of the C-linker parallel to the membrane plane (i.e. horizontally) induced a rotational movement resulting in a distinct tilting of the transmembrane helices. This movement, in turn, increased the distance between the voltage-sensing S4 domain and the surrounding transmembrane domains and led to a widening of the intracellular channel gate. In conclusion, our computational approach, combined with experimental data, thus provides a more detailed understanding of how cAMP binding is mechanically coupled over long distances to promote voltage-dependent opening of HCN channels.
Keywords: HCN1 channel; anisotropic network model; cAMP dependent gating; computational biology; cyclic AMP (cAMP); linear response theory; potassium channel; protein conformation; protein dynamic.
© 2018 Gross et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures







Similar articles
-
The HCN domain couples voltage gating and cAMP response in hyperpolarization-activated cyclic nucleotide-gated channels.Elife. 2019 Nov 26;8:e49672. doi: 10.7554/eLife.49672. Elife. 2019. PMID: 31769408 Free PMC article.
-
The HCN domain is required for HCN channel cell-surface expression and couples voltage- and cAMP-dependent gating mechanisms.J Biol Chem. 2020 Jun 12;295(24):8164-8173. doi: 10.1074/jbc.RA120.013281. Epub 2020 Apr 27. J Biol Chem. 2020. PMID: 32341127 Free PMC article.
-
A mechanism for the auto-inhibition of hyperpolarization-activated cyclic nucleotide-gated (HCN) channel opening and its relief by cAMP.J Biol Chem. 2014 Aug 8;289(32):22205-20. doi: 10.1074/jbc.M114.572164. Epub 2014 May 30. J Biol Chem. 2014. PMID: 24878962 Free PMC article.
-
The structure of the apo cAMP-binding domain of HCN4 - a stepping stone toward understanding the cAMP-dependent modulation of the hyperpolarization-activated cyclic-nucleotide-gated ion channels.FEBS J. 2018 Jun;285(12):2182-2192. doi: 10.1111/febs.14408. Epub 2018 Mar 14. FEBS J. 2018. PMID: 29444387 Review.
-
Structural and functional approaches to studying cAMP regulation of HCN channels.Biochem Soc Trans. 2021 Dec 17;49(6):2573-2579. doi: 10.1042/BST20210290. Biochem Soc Trans. 2021. PMID: 34812892 Review.
Cited by
-
Validation of the binding stoichiometry between HCN channels and their neuronal regulator TRIP8b by single molecule measurements.Front Physiol. 2022 Sep 26;13:998176. doi: 10.3389/fphys.2022.998176. eCollection 2022. Front Physiol. 2022. PMID: 36225302 Free PMC article.
-
Review: HCN Channels in the Heart.Curr Cardiol Rev. 2022;18(4):e040222200836. doi: 10.2174/1573403X18666220204142436. Curr Cardiol Rev. 2022. PMID: 35125083 Free PMC article. Review.
-
Weak Cation Selectivity in HCN Channels Results From K+-Mediated Release of Na+ From Selectivity Filter Binding Sites.Function (Oxf). 2022 Apr 22;3(3):zqac019. doi: 10.1093/function/zqac019. eCollection 2022. Function (Oxf). 2022. PMID: 36156894 Free PMC article.
-
Seizures, behavioral deficits, and adverse drug responses in two new genetic mouse models of HCN1 epileptic encephalopathy.Elife. 2022 Aug 16;11:e70826. doi: 10.7554/eLife.70826. Elife. 2022. PMID: 35972069 Free PMC article.
-
The HCN domain couples voltage gating and cAMP response in hyperpolarization-activated cyclic nucleotide-gated channels.Elife. 2019 Nov 26;8:e49672. doi: 10.7554/eLife.49672. Elife. 2019. PMID: 31769408 Free PMC article.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
