

mTORC1 Controls Phase Separation and the Biophysical Properties of the Cytoplasm by Tuning Crowding
- PMID: 29937223
- PMCID: PMC10080728
- DOI: 10.1016/j.cell.2018.05.042
mTORC1 Controls Phase Separation and the Biophysical Properties of the Cytoplasm by Tuning Crowding
Abstract
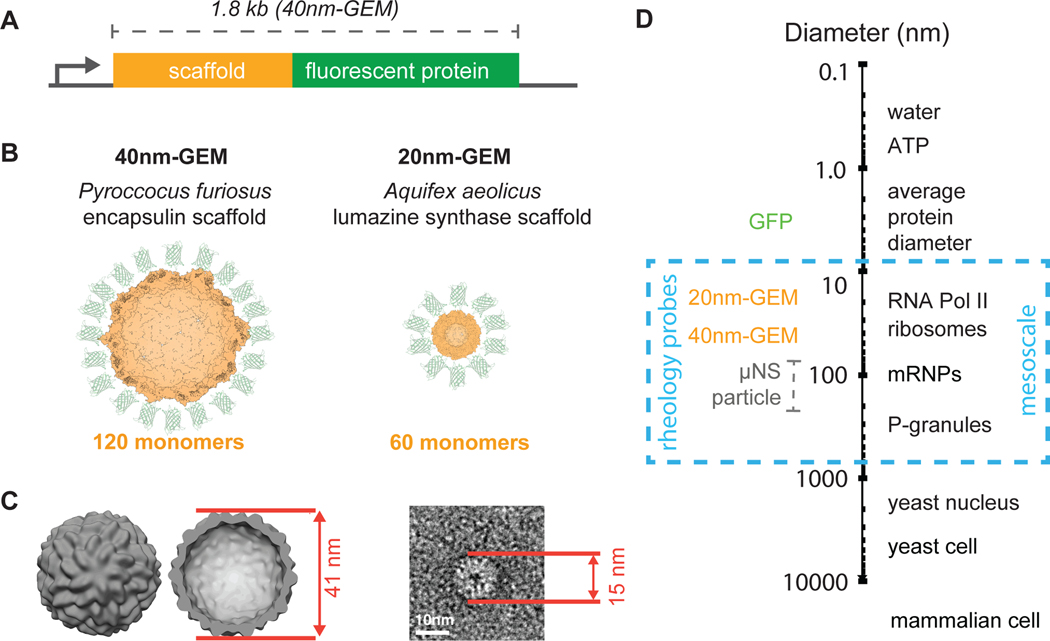
Macromolecular crowding has a profound impact on reaction rates and the physical properties of the cell interior, but the mechanisms that regulate crowding are poorly understood. We developed genetically encoded multimeric nanoparticles (GEMs) to dissect these mechanisms. GEMs are homomultimeric scaffolds fused to a fluorescent protein that self-assemble into bright, stable particles of defined size and shape. By combining tracking of GEMs with genetic and pharmacological approaches, we discovered that the mTORC1 pathway can modulate the effective diffusion coefficient of particles ≥20 nm in diameter more than 2-fold by tuning ribosome concentration, without any discernable effect on the motion of molecules ≤5 nm. This change in ribosome concentration affected phase separation both in vitro and in vivo. Together, these results establish a role for mTORC1 in controlling both the mesoscale biophysical properties of the cytoplasm and biomolecular condensation.
Keywords: biophysics; cytoplasm; electron tomography; mTORC1; microrheology; molecular crowding; nanoparticles; phase separation; ribosomes; systems biology.
Copyright © 2018 Elsevier Inc. All rights reserved.
Figures






Comment in
-
Translating the Physical Code of Life.Cell. 2018 Jul 12;174(2):253-255. doi: 10.1016/j.cell.2018.06.050. Cell. 2018. PMID: 30007414 Free PMC article.
-
Crowd control.Nat Chem Biol. 2018 Aug;14(8):745. doi: 10.1038/s41589-018-0109-1. Nat Chem Biol. 2018. PMID: 30018416 No abstract available.
References
-
- Akita F, Chong KT, Tanaka H, Yamashita E, Miyazaki N, Nakaishi Y, Suzuki M, Namba K, Ono Y, Tsukihara T, et al. (2007). The crystal structure of a virus-like particle from the hyperthermophilic archaeon Pyrococcus furiosus provides insight into the evolution of viruses. J Mol Biol 368, 1469–1483. - PubMed
-
- Asano S, Engel BD, and Baumeister W. (2016). In Situ Cryo-Electron Tomography: A Post-Reductionist Approach to Structural Biology (Academic Press; ). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
