

Vascular Compartmentalization of Functional Hyperemia from the Synapse to the Pia
- PMID: 29937277
- PMCID: PMC6069674
- DOI: 10.1016/j.neuron.2018.06.012
Vascular Compartmentalization of Functional Hyperemia from the Synapse to the Pia
Erratum in
-
Vascular Compartmentalization of Functional Hyperemia from the Synapse to the Pia.Neuron. 2019 Feb 20;101(4):762. doi: 10.1016/j.neuron.2019.01.060. Neuron. 2019. PMID: 30790540 Free PMC article. No abstract available.
Abstract
Functional hyperemia, a regional increase of blood flow triggered by local neural activation, is used to map brain activity in health and disease. However, the spatial-temporal dynamics of functional hyperemia remain unclear. Two-photon imaging of the entire vascular arbor in NG2-creERT2;GCaMP6f mice shows that local synaptic activation, measured via oligodendrocyte precursor cell (OPC) Ca2+ signaling, generates a synchronous Ca2+ drop in pericytes and smooth muscle cells (SMCs) enwrapping all upstream vessels feeding the activated synapses. Surprisingly, the onset timing, direction, and amplitude of vessel diameter and blood velocity changes vary dramatically from juxta-synaptic capillaries back to the pial arteriole. These results establish a precise spatial-temporal sequence of vascular changes triggered by neural activity and essential for the interpretation of blood-flow-based imaging techniques such as BOLD-fMRI.
Keywords: CBF; anesthetized; astrocyte; awake; blood-brain barrier; calcium; endothelium; functional imaging; gap junction; glia; glutamate; hyperpolarization; in vivo; microvascular; neuron; neurovascular coupling; neurovascular unit; odor.
Copyright © 2018 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
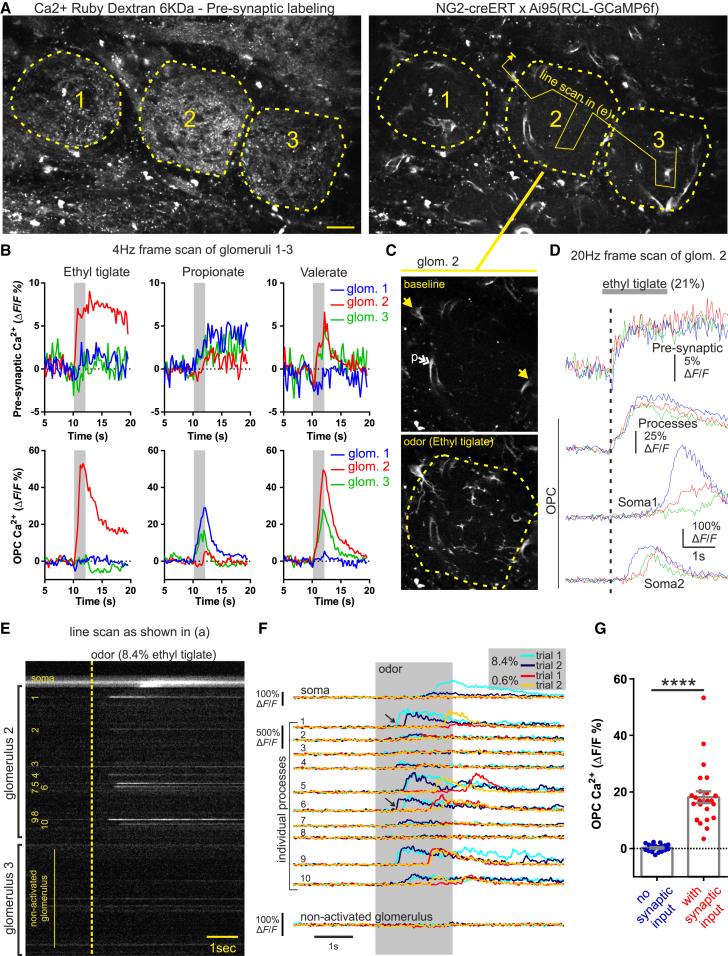
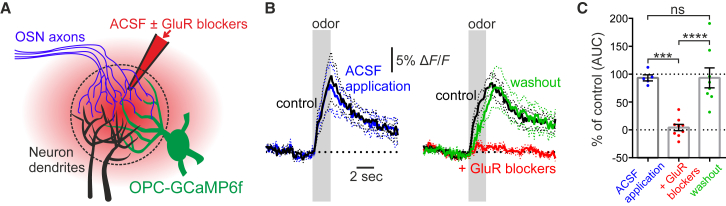
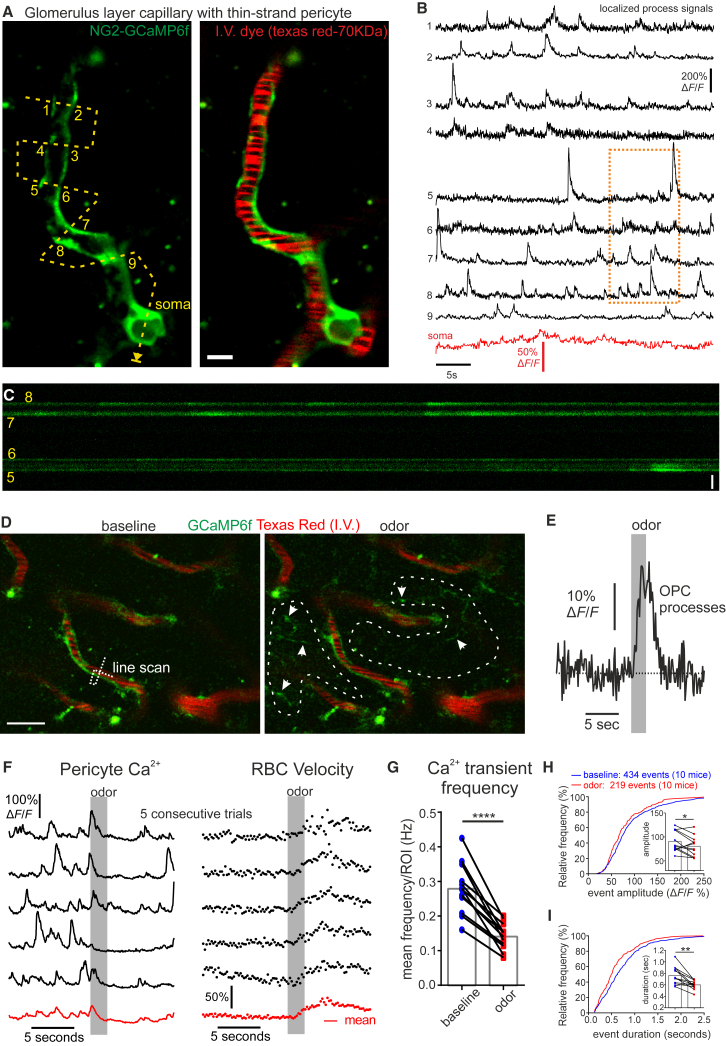
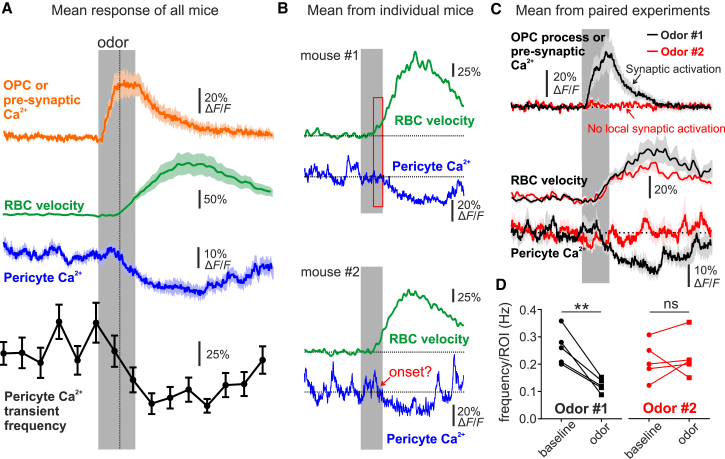
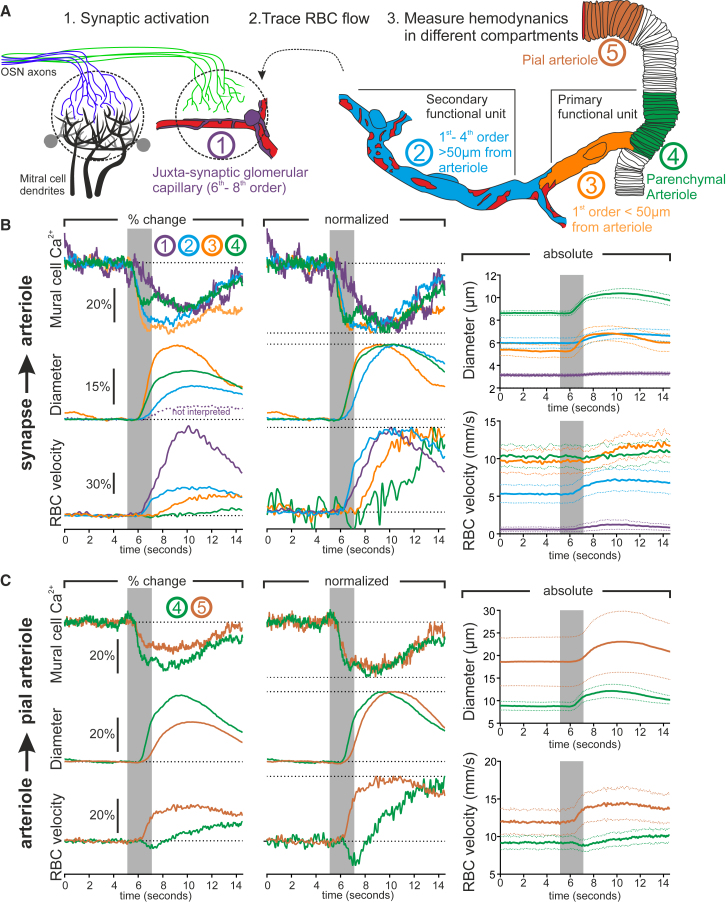
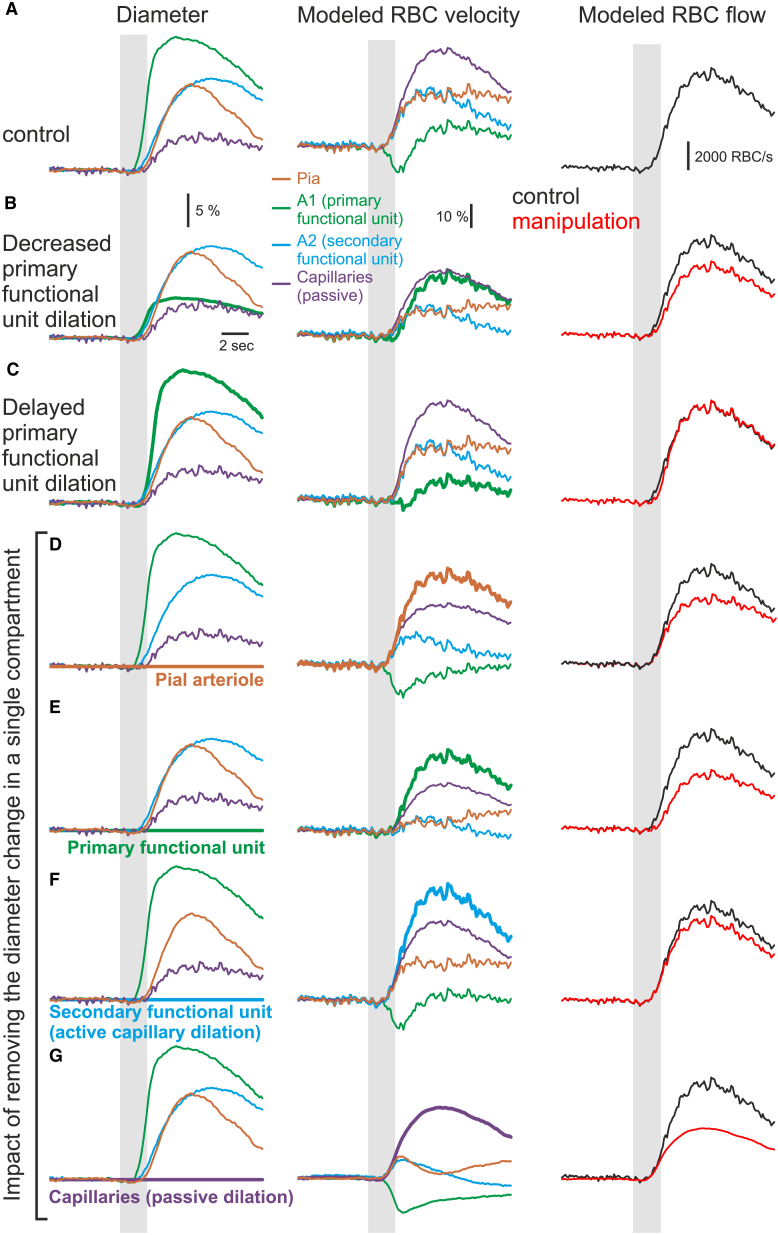
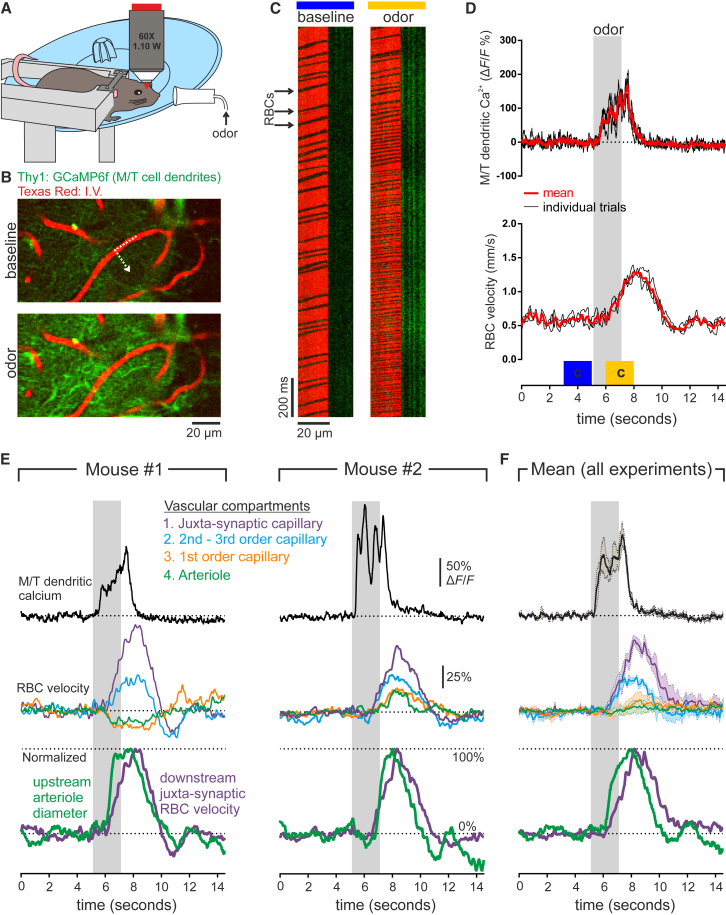
Comment in
-
Keeping the Brain Well Fed: The Role of Capillaries and Arterioles in Orchestrating Functional Hyperemia.Neuron. 2018 Jul 25;99(2):248-250. doi: 10.1016/j.neuron.2018.07.011. Neuron. 2018. PMID: 30048612 Free PMC article.
References
-
- Bergles D.E., Roberts J.D., Somogyi P., Jahr C.E. Glutamatergic synapses on oligodendrocyte precursor cells in the hippocampus. Nature. 2000;405:187–191. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
