

Review
doi: 10.1387/ijdb.170341fw.
Homeostasis, regeneration and tumour formation in the mammalian epidermis
Affiliations
- PMID: 29938768
- PMCID: PMC6103439
- DOI: 10.1387/ijdb.170341fw
Item in Clipboard
Review
Homeostasis, regeneration and tumour formation in the mammalian epidermis
Int J Dev Biol.
2018.
Abstract
The epidermis is the outer covering of the skin and provides a protective interface between the body and the environment. It is well established that the epidermis is maintained by stem cells that self-renew and generate differentiated cells. In this review, we discuss how recent technological advances, including single cell transcriptomics and in vivo imaging, have provided new insights into the nature and plasticity of the stem cell compartment and the differing roles of stem cells in homeostasis, wound repair and cancer.
Figures
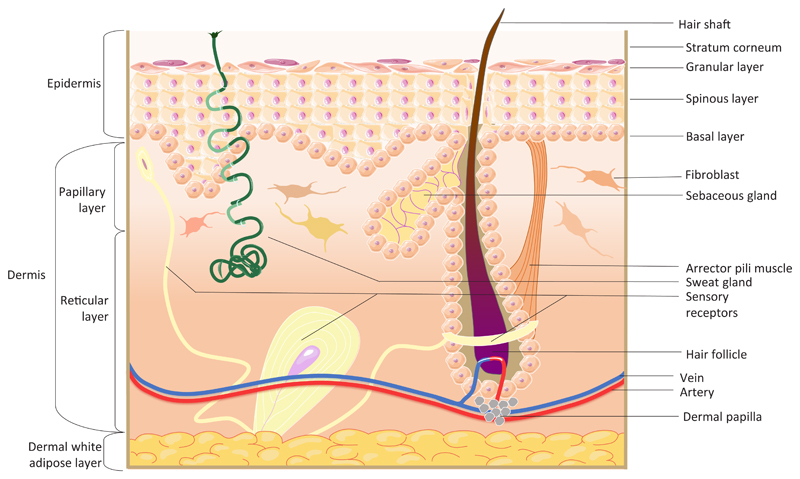
Skin consists of two main layers, the epidermis and dermis. The epidermis between hair follicles is known as the interfollicular epidermis (IFE) which consists of keratinocytes arranged into layers: basal, spinous, granular and stratum corneum. The epidermis also forms adnexal structures, such as hair follicles, sebaceous glands and sweat glands. A basement membrane separates the epidermis from the dermis. The dermis consists of several layers: papillary, reticular and hypodermis/white adipose tissue. The dermis also contains blood vessels, sensory nerves, arrector pili muscles (which control pilo-erection) and dermal papillae, clusters of fibroblasts at the base of hair follicles that regulate the hair growth cycle.
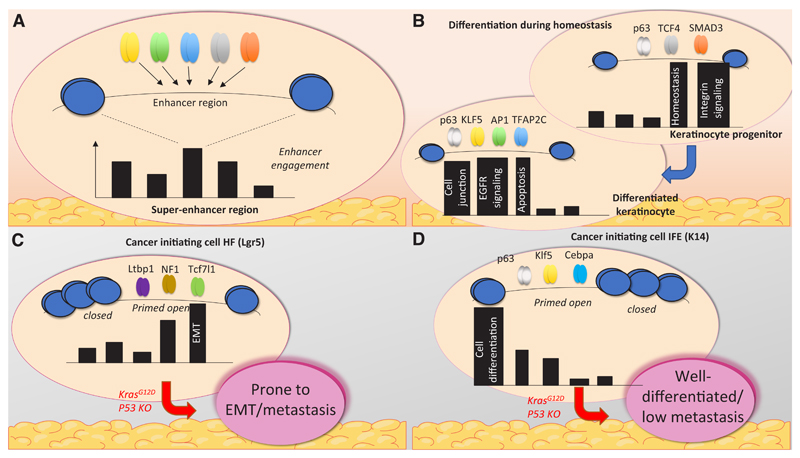
Super-enhancers are extensive regions within the genome containing multiple TF-recruiting sequences, i.e. enhancers. (A) Super-enhancer usage changes during differentiation of keratinocytes. Recruitment of control regions, involved in homeostasis and integrin signalling, is reduced, and replaced by areas responsible for genes involved in cell-cell junctions, apoptosis and EGFR signalling. (B) Enhancer recruitment differs in various types of cells, such as HF or IFE cells. In tumours, these enhancers determine the properties of the tumour by engagement of different genes. For example, if the cancer-originating cell is of HF origin, genes that are primed open will more likely cause metastasis (C). On the contrary, IFE cells contain enhancers primed towards cell differentiation, and hence tumours are better differentiated and rarely metastasise (D).
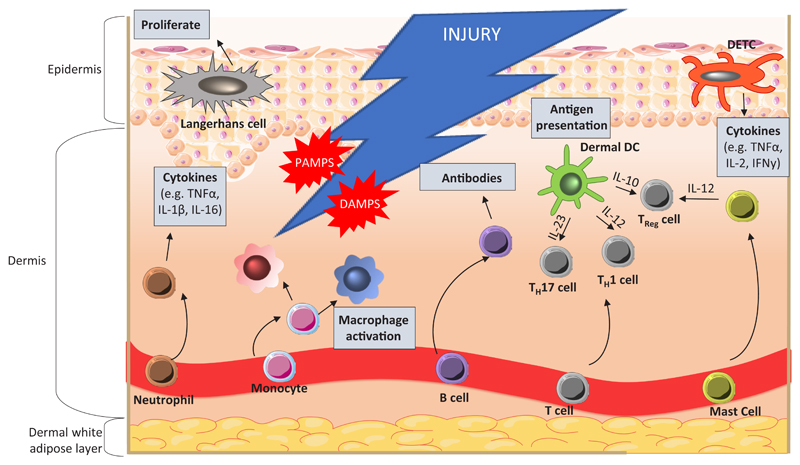
During injury, resident skin cells, including keratinocytes, macrophages, and dendritic cells, are exposed to danger signals from host cells, DAMPs (damage associated molecular patterns), invading microorganisms and PAMPs (pathogen associated molecular patterns). Neutrophil invasion is required initially for phagocytosis and cytokine secretion, to amplify the immune response. Infiltration and maturation of monocytes into a spectrum of macrophage phenotypes occurs to support inflammation. Dermal dendritic cells (DCs) aid in T cell polarisation to TH1, TH17 and Treg through presentation of antigens and release of cytokines. Langerhans cells are involved in immune surveillance and are the first-line of defence in epidermal wounds. Upon inflammation, cells become activated and promote re-epithelialisation through induction of keratinocyte proliferation and differentiation. DETCs are the only resident T cells in mouse epidermis and are crucial for immune surveillance and homeostasis. Signals from damaged keratinocytes activate DETCs, which regulate further immune responses. Mast cells and dermal DCs establish the balance between pro- and anti-inflammatory responses through secretion of specific cytokines. An adaptive immune response occurs after the initial inflammation, and involves B and T cells. B cells produce antibodies against invading pathogens whilst T cells secrete cytokines and growth factors.
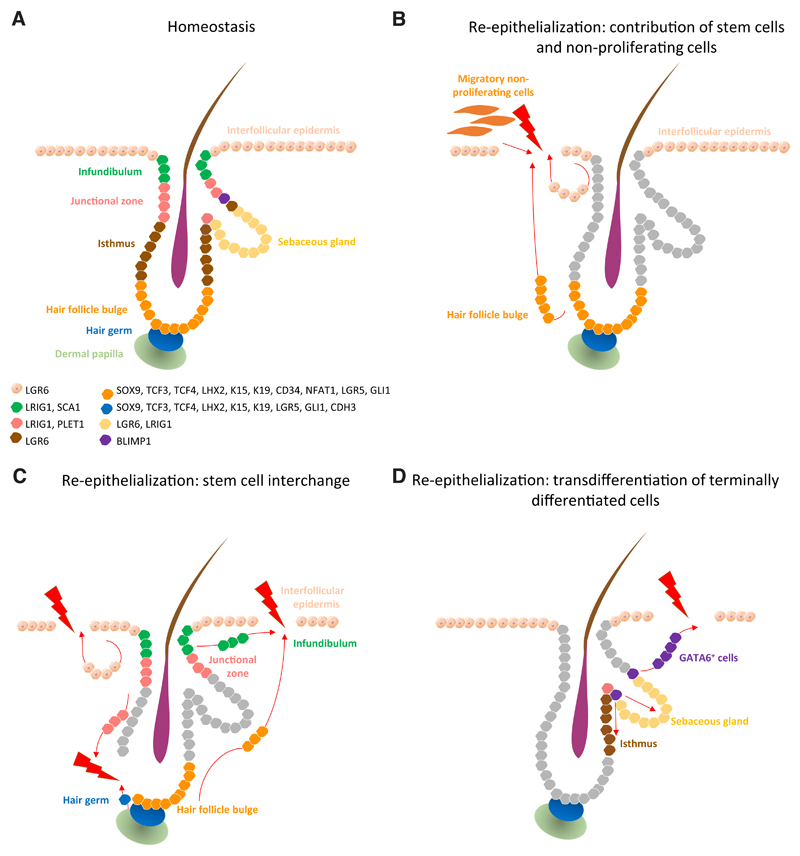
Epidermal stem cell compartments that maintain skin homeostasis and their associated markers (A). Re-epithelialization upon injury occurs via several paths: contribution of the proliferative hub (IFE hair- follicle stem cells and their progeny) and non-proliferative migratory cells (at the leading edge) to the initial stages of re-epithelialization (B). When stem cell compartments from the IFE, infundibulum, junctional zone and hair follicle bulge and germ exhibit plasticity, they contribute to the replenishment of stem cells lost on wounding (C). Terminally differentiated cells such as GATA6+ cells de-differentiate and contribute to re- epithelialization of damaged IFE and re-populate the sebaceous gland and lower hair follicle during wound healing (D).
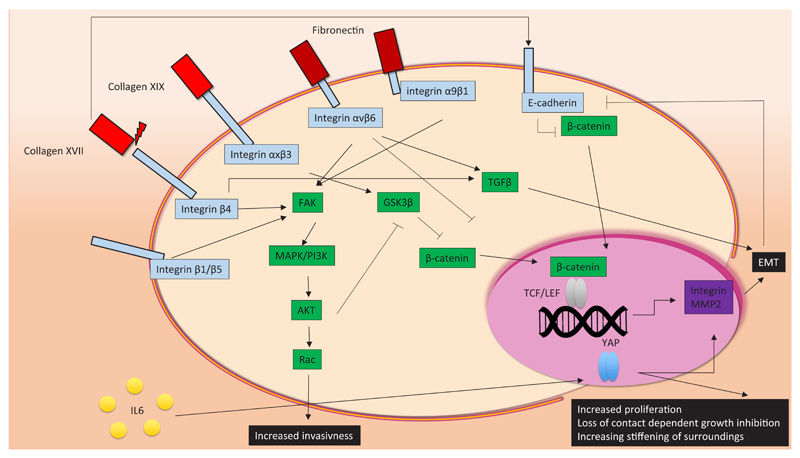
Components of the extracellular matrix (ECM), such as collagens and fibronectin, interact with cell surface integrins and cadherins. Depending on which surface molecule they interact with, the cancer hallmarks (uncontrolled proliferation, invasiveness and epithelial-to-mesenchymal transition (EMT)) can either be activated or repressed. Activation of the MAPK/PI3K pathway, mediated by integrin stimulation, facilitates nuclear translocation of β-catenin, resulting in EMT and increased invasiveness. EMT is also induced by TGFβ signalling, through collagen- and fibronectin-integrin associations. Other stimuli, including secreted cytokines, can contribute to cell transformation. For example, YAP signalling is activated by IL-5, in turn causing increased proliferation and loss of contact-dependent growth inhibition, which is typical behaviour associated with cancer cells.
References
-
- Arwert EN, Hoste E, Watt FM. Epithelial stem cells, wound healing and cancer. Nat Rev Cancer. 2012;12:170–180. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
