

Lipin-1 regulates Bnip3-mediated mitophagy in glycolytic muscle
- PMID: 29939786
- PMCID: PMC6219840
- DOI: 10.1096/fj.201800374
Lipin-1 regulates Bnip3-mediated mitophagy in glycolytic muscle
Abstract
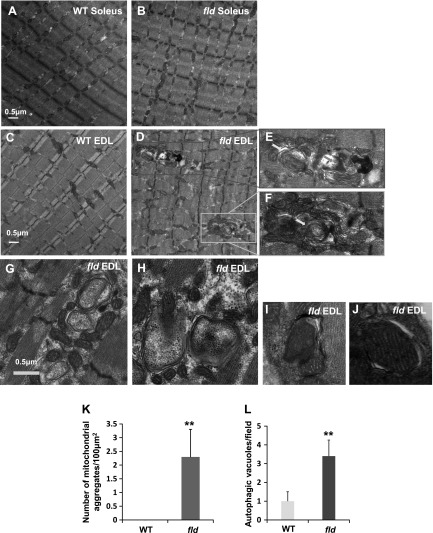
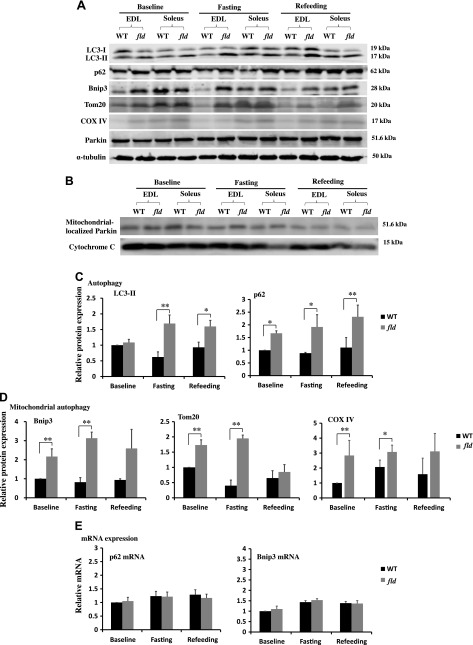
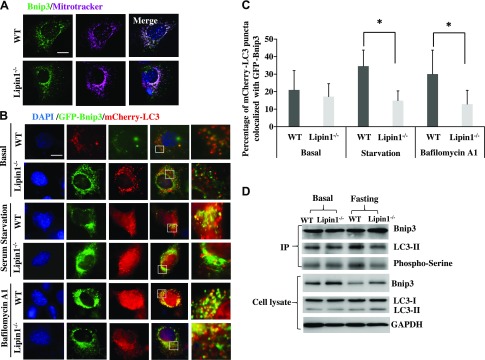
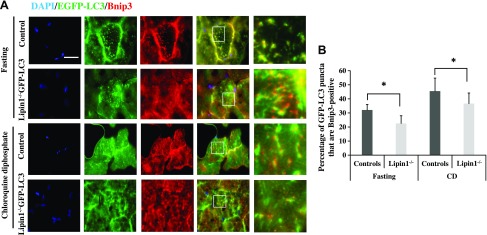
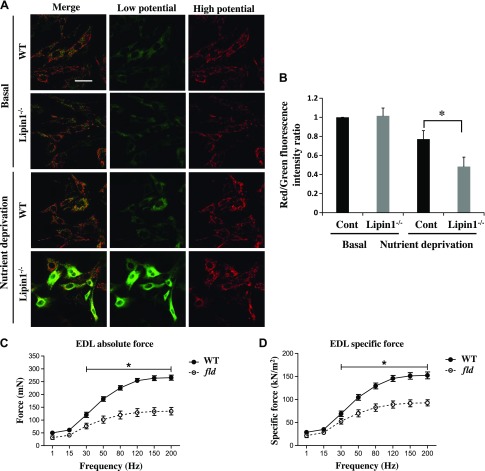
Autophagy of mitochondria (mitophagy) is essential for maintaining muscle mass and healthy skeletal muscle. Patients with heritable phosphatidic acid phosphatase lipin-1-null mutations present with severe rhabdomyolysis and muscle atrophy in glycolytic muscle fibers, which are accompanied with mitochondrial aggregates and reduced mitochondrial cytochrome c oxidase activity. However, the underlying mechanisms leading to muscle atrophy as a result of lipin-1 deficiency are still not clear. In this study, we found that lipin-1 deficiency in mice is associated with a marked accumulation of abnormal mitochondria and autophagic vacuoles in glycolytic muscle fibers. Our studies using lipin-1-deficient myoblasts suggest that lipin-1 participates in B-cell leukemia (BCL)-2 adenovirus E1B 19 kDa protein-interacting protein 3 (Bnip3)-regulated mitophagy by interacting with microtubule-associated protein 1A/1B-light chain (LC)3, which is an important step in the recruitment of mitochondria to nascent autophagosomes. The requirement of lipin-1 for Bnip3-mediated mitophagy was further verified in vivo in lipin-1-deficient green fluorescent protein-LC3 transgenic mice (lipin-1-/--GFP-LC3). Finally, we showed that lipin-1 deficiency in mice resulted in defective mitochondrial adaptation to starvation-induced metabolic stress and impaired contractile muscle force in glycolytic muscle fibers. In summary, our study suggests that deregulated mitophagy arising from lipin-1 deficiency is associated with impaired muscle function and may contribute to muscle rhabdomyolysis in humans.-Alshudukhi, A. A., Zhu, J., Huang, D., Jama, A., Smith, J. D., Wang, Q. J., Esser, K. A., Ren, H. Lipin-1 regulates Bnip3-mediated mitophagy in glycolytic muscle.
Keywords: LC3; contractile force; fld; mitochondrial autophagy; rhabdomyolysis.
Conflict of interest statement
The authors thank Dr. Noboro Mizushima (University of Tokyo, Tokyo, Japan) for the GFP-LC3 mice, Dr. Wen-Xing Ding (University of Kansas Medical School, Kansas City, KS, USA) for the GFP-Bnip3 plasmid, and Dr. Feng Zhang (McGovern Institute for Brian Research, Massachusetts Institute of Technology, Cambridge, MA, USA) for pSpCas9 (BB)-2A-Puro (PX495) v.2.0 (plasmid 62988; Addgene, Cambridge, MA, USA). This work was supported by startup funds from Wright State University (to H.R.), U.S. National Institutes of Health Center of Biomedical Research Excellence on Obesity and Cardiovascular Diseases Grant P20 GM103527-06, and Beginning Grant-in-Aid 11BGIA7710059 and Scientist Development Grant 12SDG12050697 from the American Heart Association (to H.R.). The authors declare no conflicts of interest.
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
