

Aspartate is an endogenous metabolic limitation for tumour growth
- PMID: 29941931
- PMCID: PMC6051729
- DOI: 10.1038/s41556-018-0125-0
Aspartate is an endogenous metabolic limitation for tumour growth
Abstract
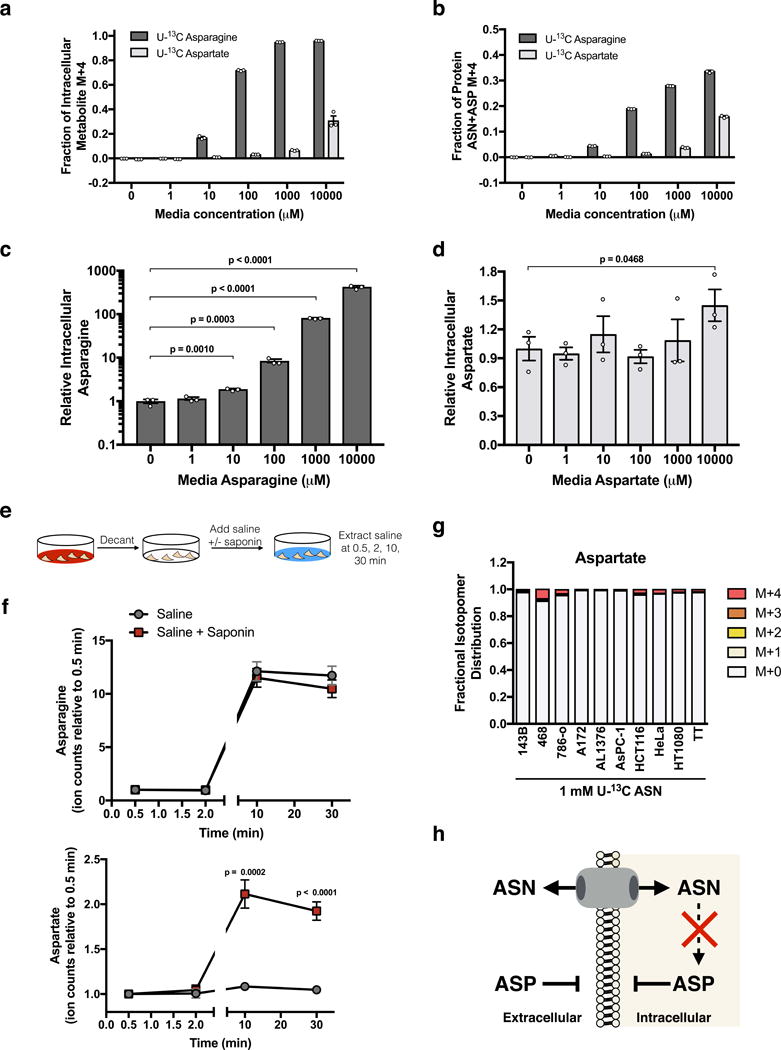
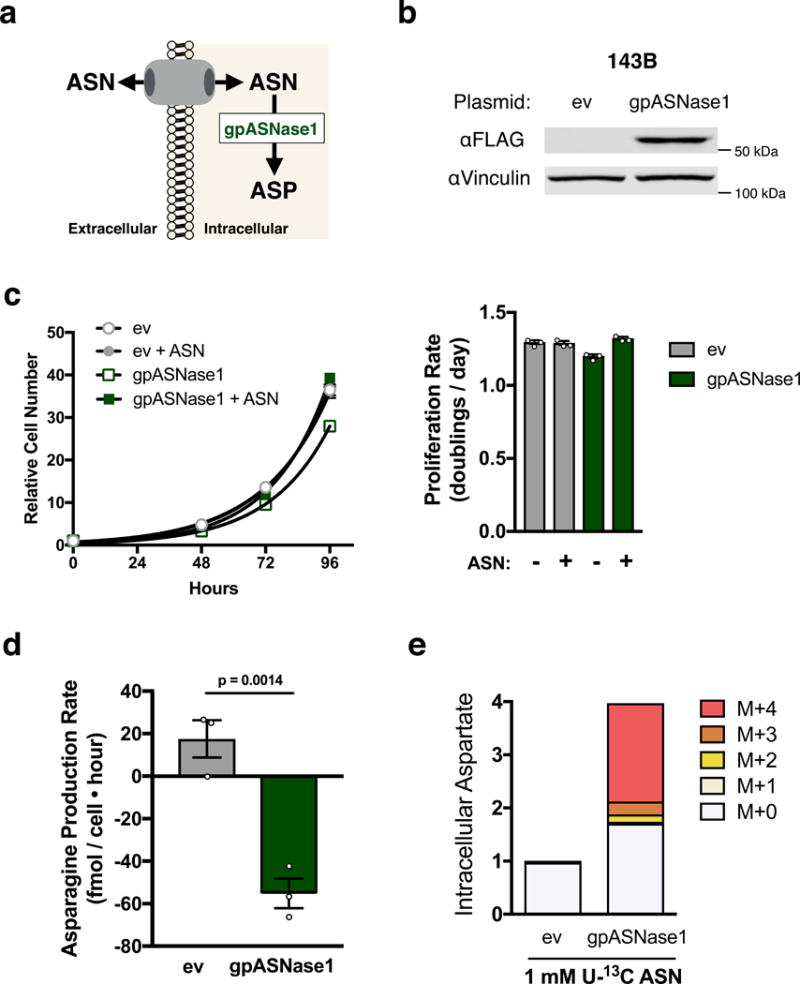
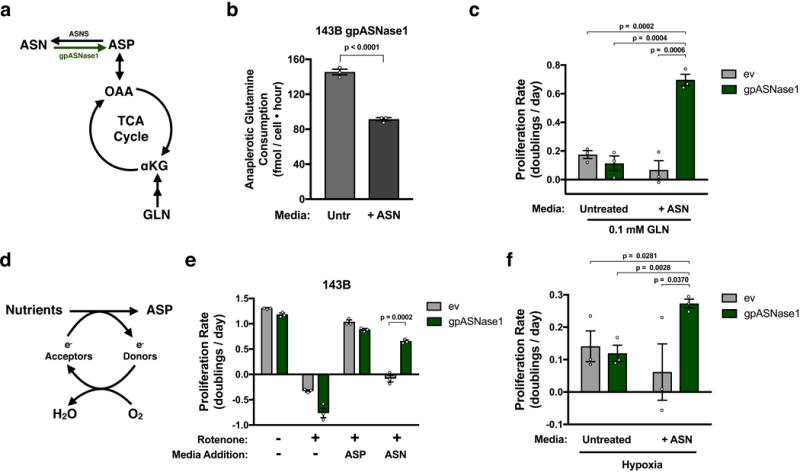
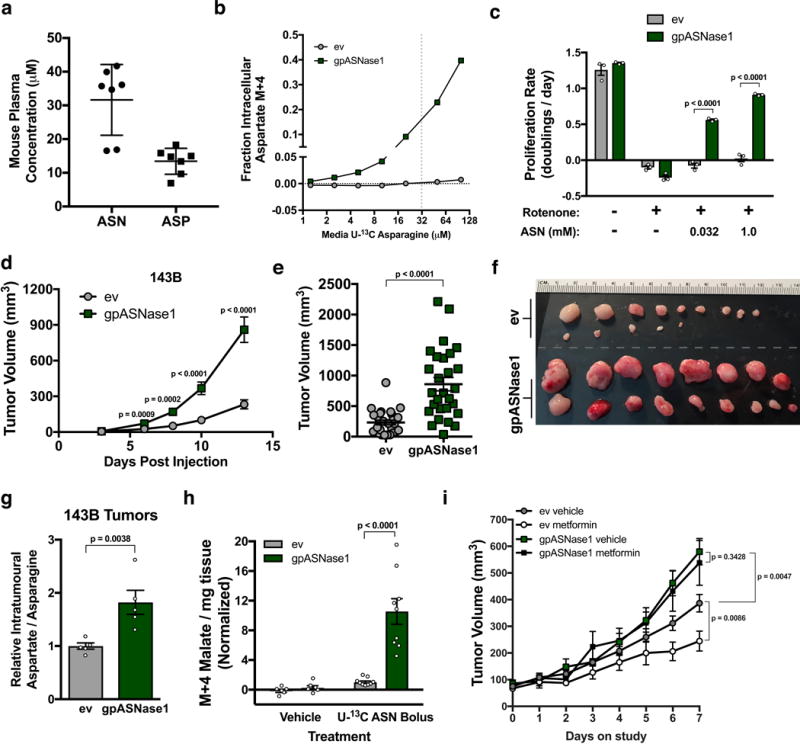
Defining the metabolic limitations of tumour growth will help to develop cancer therapies1. Cancer cells proliferate slower in tumours than in standard culture conditions, indicating that a metabolic limitation may restrict cell proliferation in vivo. Aspartate synthesis can limit cancer cell proliferation when respiration is impaired2-4; however, whether acquiring aspartate is endogenously limiting for tumour growth is unknown. We confirm that aspartate has poor cell permeability, which prevents environmental acquisition, whereas the related amino acid asparagine is available to cells in tumours, but cancer cells lack asparaginase activity to convert asparagine to aspartate. Heterologous expression of guinea pig asparaginase 1 (gpASNase1), an enzyme that produces aspartate from asparagine5, confers the ability to use asparagine to supply intracellular aspartate to cancer cells in vivo. Tumours expressing gpASNase1 grow at a faster rate, indicating that aspartate acquisition is an endogenous metabolic limitation for the growth of some tumours. Tumours expressing gpASNase1 are also refractory to the growth suppressive effects of metformin, suggesting that metformin inhibits tumour growth by depleting aspartate. These findings suggest that therapeutic aspartate suppression could be effective to treat cancer.
Figures
Comment in
-
Grasping for aspartate in tumour metabolism.Nat Cell Biol. 2018 Jul;20(7):738-739. doi: 10.1038/s41556-018-0137-9. Nat Cell Biol. 2018. PMID: 29941932 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
