

Towards a gene regulatory network shaping the fins of the Princess cichlid
- PMID: 29942008
- PMCID: PMC6018552
- DOI: 10.1038/s41598-018-27977-y
Towards a gene regulatory network shaping the fins of the Princess cichlid
Abstract
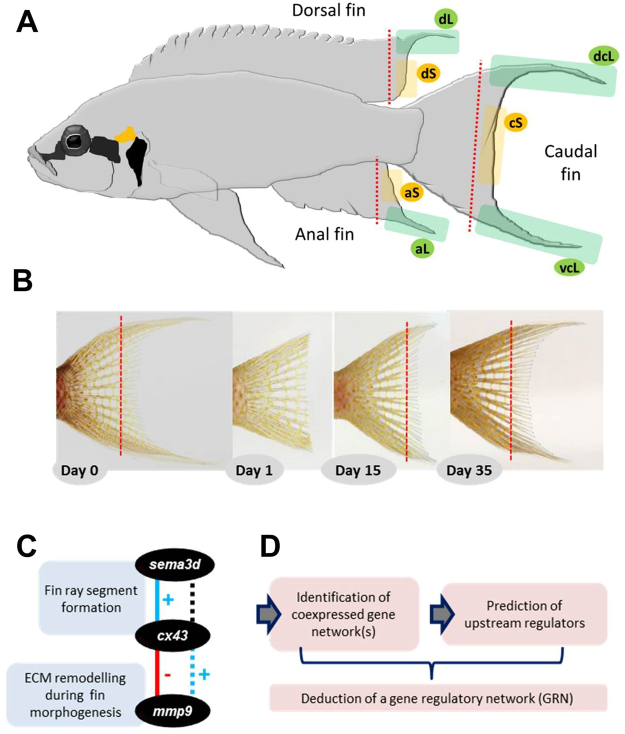
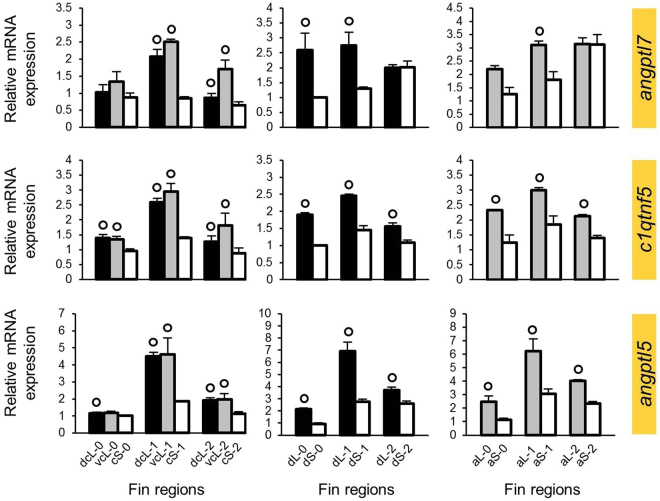
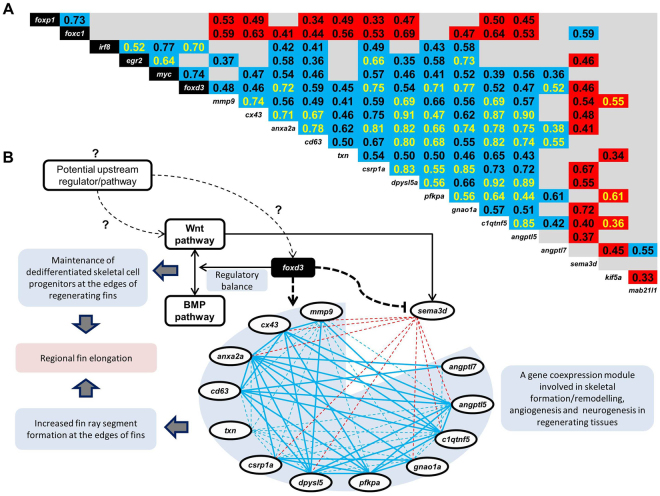
Variation in fin shape and size contributes to the outstanding morphological diversity of teleost fishes, but the regulation of fin growth has not yet been studied extensively outside the zebrafish model. A previous gene expression study addressing the ornamental elongations of unpaired fins in the African cichlid fish Neolamprologus brichardi identified three genes (cx43, mmp9 and sema3d) with strong and consistent expression differences between short and elongated fin regions. Remarkably, the expression patterns of these genes were not consistent with inferences on their regulatory interactions in zebrafish. Here, we identify a gene expression network (GRN) comprising cx43, mmp9, and possibly also sema3d by a stepwise approach of identifying co-expression modules and predicting their upstream regulators. Among the transcription factors (TFs) predicted as potential upstream regulators of 11 co-expressed genes, six TFs (foxc1, foxp1, foxd3, myc, egr2, irf8) showed expression patterns consistent with their cooperative transcriptional regulation of the gene network. Some of these TFs have already been implicated in teleost fish fin regeneration and formation. We particularly discuss the potential function of foxd3 as driver of the network and its role in the unexpected gene expression correlations observed in N. brichardi.
Conflict of interest statement
The authors declare no competing interests.
Figures



References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
