

Tight Turns of Outer Membrane Proteins: An Analysis of Sequence, Structure, and Hydrogen Bonding
- PMID: 29944853
- PMCID: PMC6110081
- DOI: 10.1016/j.jmb.2018.06.013
Tight Turns of Outer Membrane Proteins: An Analysis of Sequence, Structure, and Hydrogen Bonding
Abstract
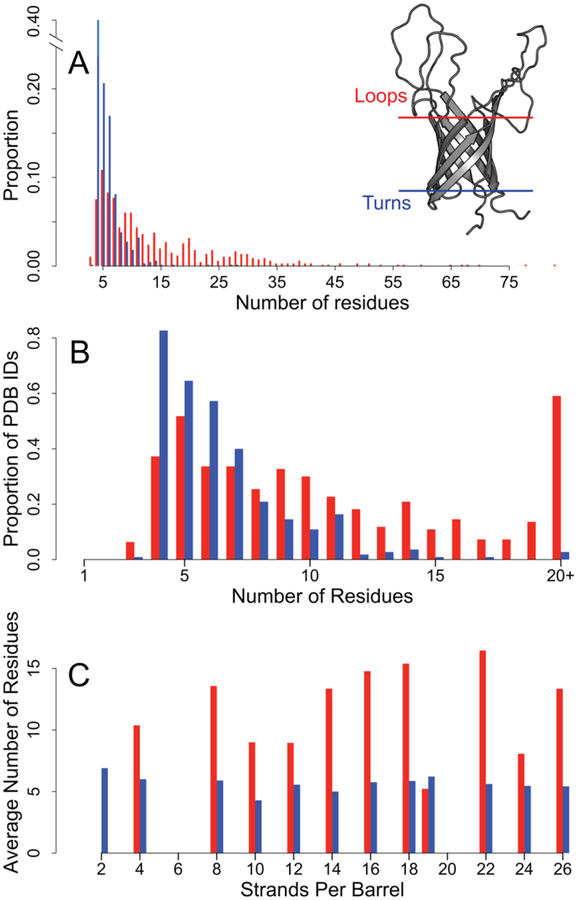
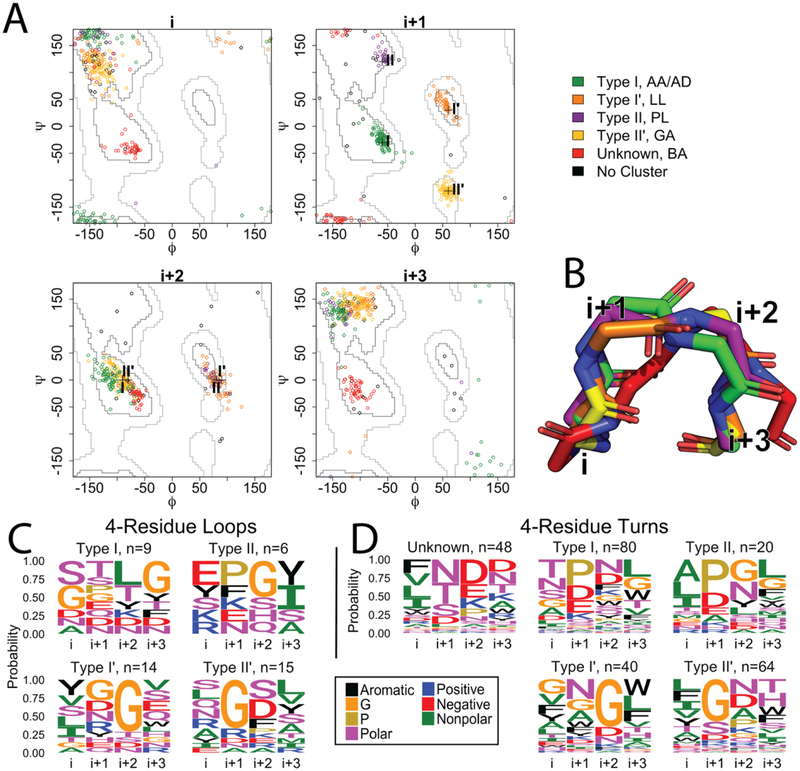
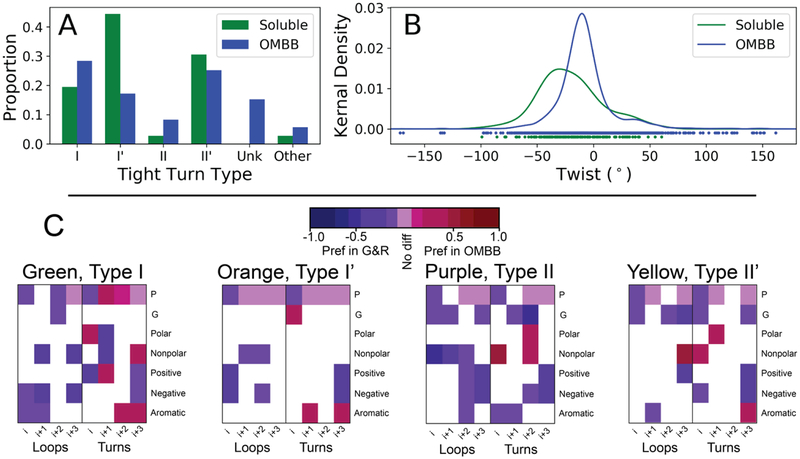
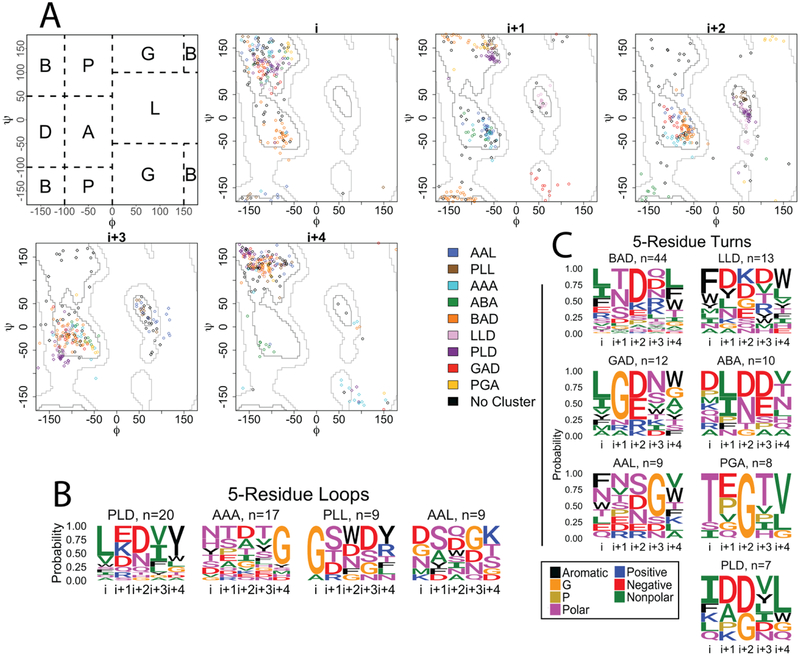
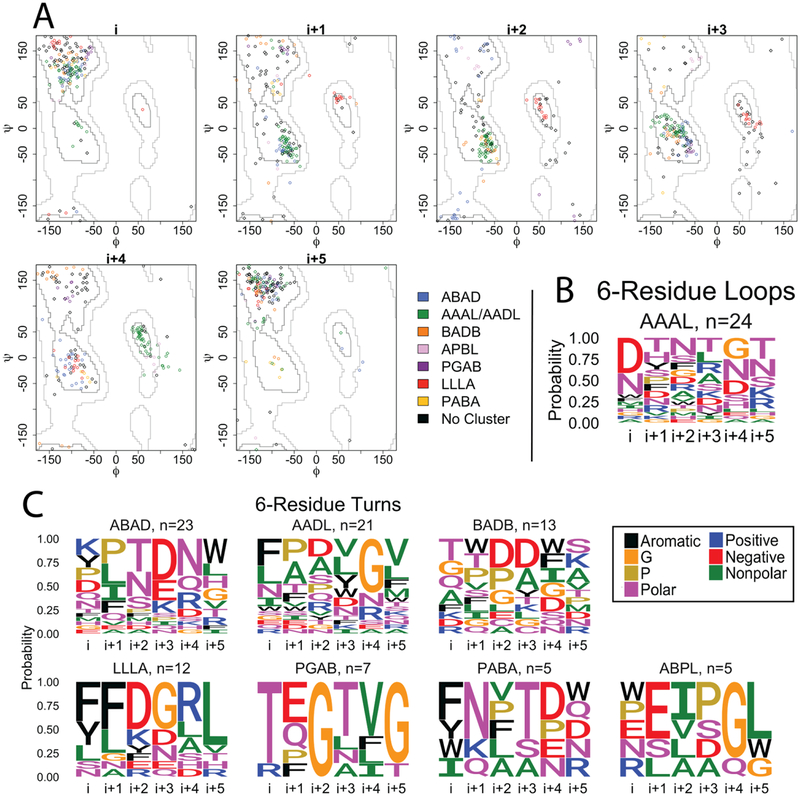
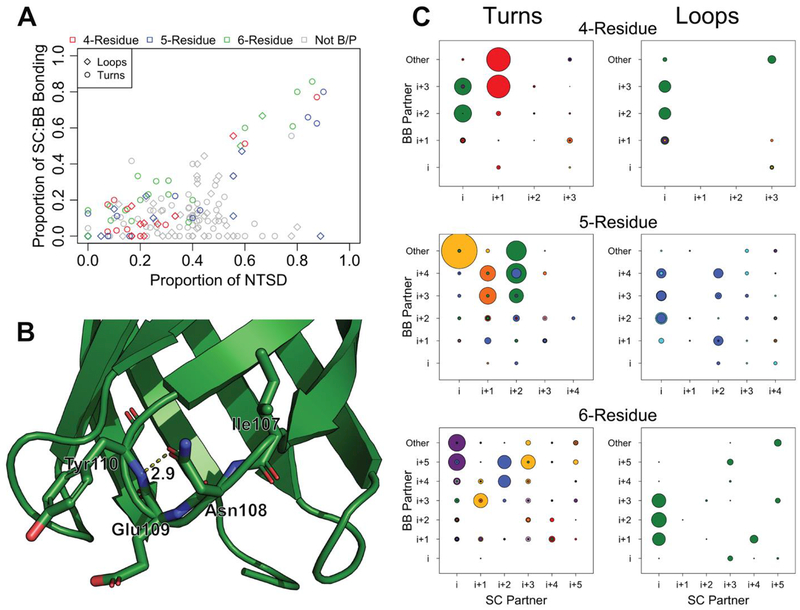
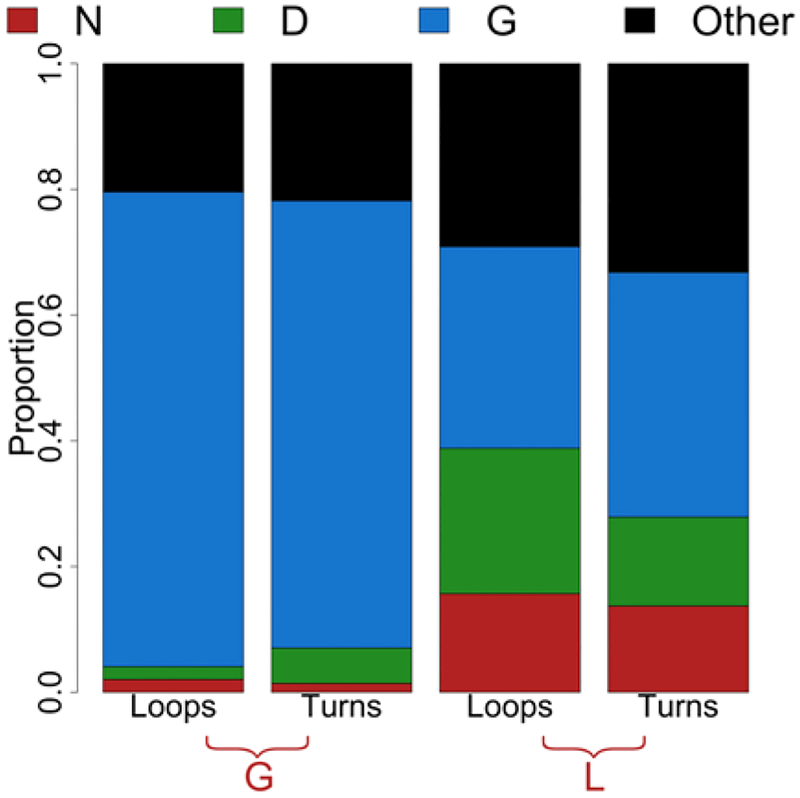
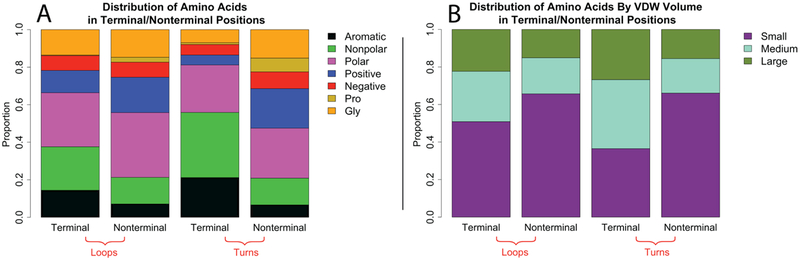
As a structural class, tight turns can control molecular recognition, enzymatic activity, and nucleation of folding. They have been extensively characterized in soluble proteins but have not been characterized in outer membrane proteins (OMPs), where they also support critical functions. We clustered the 4 to 6 residue tight turns of 110 OMPs to characterize the phi/psi angles, sequence, and hydrogen bonding of these structures. We find significant differences between reports of soluble protein tight turns and OMP tight turns. Since OMP strands are less twisted than soluble strands, they favor different turn structures types. Moreover, the membrane localization of OMPs yields different sequence hallmarks for their tight turns relative to soluble protein turns. We also characterize the differences in phi/psi angles, sequence, and hydrogen bonding between OMP extracellular loops and OMP periplasmic turns. As previously noted, the extracellular loops tend to be much longer than the periplasmic turns. We find that this difference in length is due to the broader distribution of lengths of the extracellular loops not a large difference in the median length. Extracellular loops also tend to have more charged residues as predicted by the charge-out rule. Finally, in all OMP tight turns, hydrogen bonding between the side chain and backbone 2 to 4 residues away from that side chain plays an important role. These bonds preferentially use an Asp, Asn, Ser, or Thr residue in a beta or pro phi/psi conformation. We anticipate that this study will be applicable to future design and structure prediction of OMPs.
Keywords: beta barrels/β-barrels; loops; reverse turns; α-turns; β-turns.
Copyright © 2018 Elsevier Ltd. All rights reserved.
Figures
References
-
- Guruprasad K, Rajkumar S. Beta and gamma-turns in proteins revisited: A new set of amino acid turn-type dependent positional preferences and potentials. J Biosci. 2000;25:143–56. - PubMed
-
- Chou K-C. Prediction of Tight Turns and Their Types in Proteins. Anal Biochem. 2000;286:1–16. - PubMed
-
- Rose GD, Gierasch LM, Smith JA. Turns in peptides and proteins. Adv Protein Chem. 1985;37:1–109. - PubMed
-
- Kee KS, Jois SD. Design of beta-turn based therapeutic agents. Current pharmaceutical design. 2003;9:1209–24. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
