

The Initial Response of a Eukaryotic Replisome to DNA Damage
- PMID: 29944888
- PMCID: PMC6024075
- DOI: 10.1016/j.molcel.2018.04.022
The Initial Response of a Eukaryotic Replisome to DNA Damage
Abstract
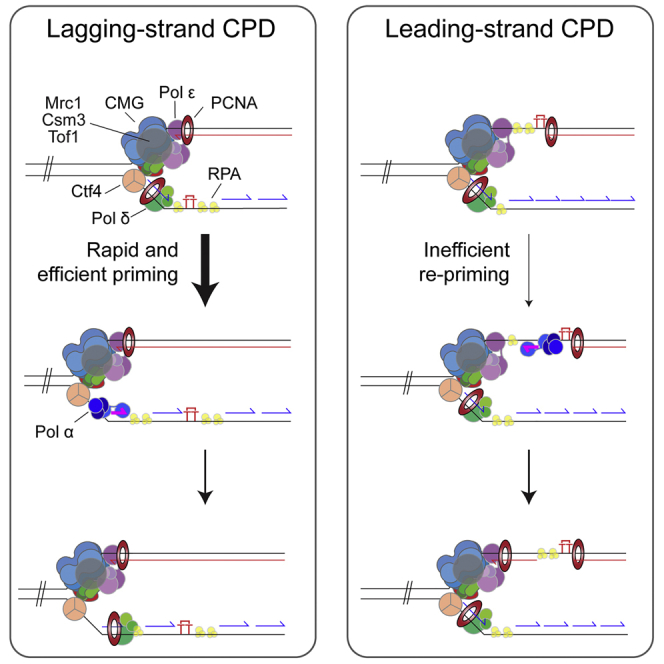
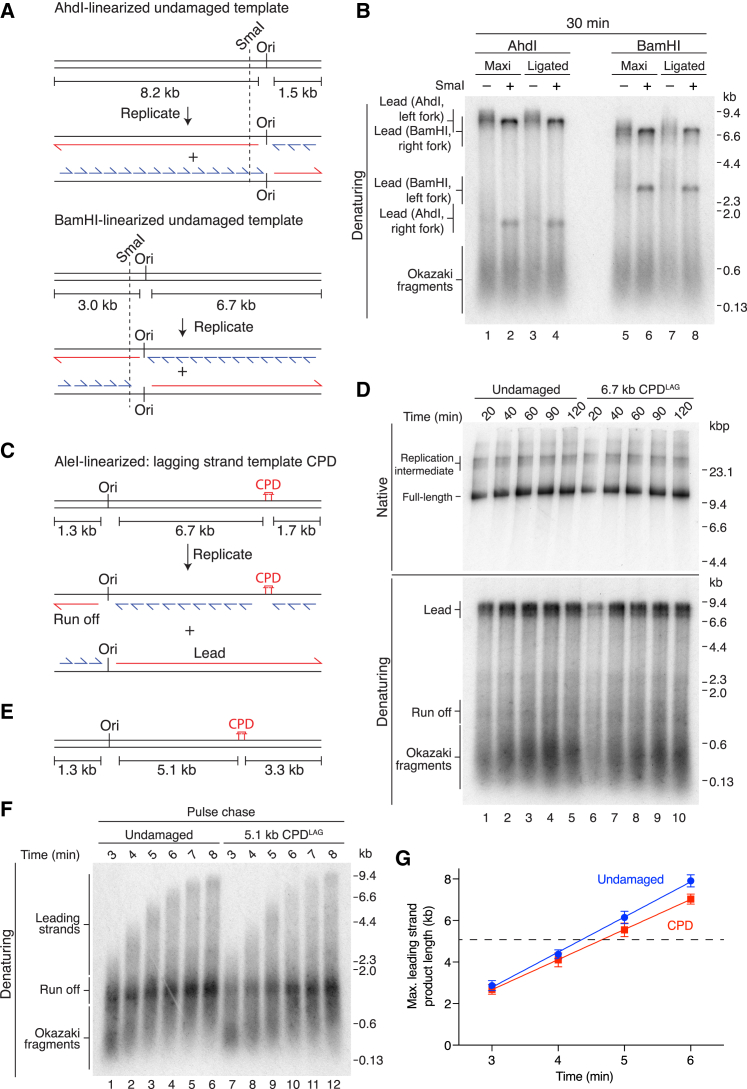
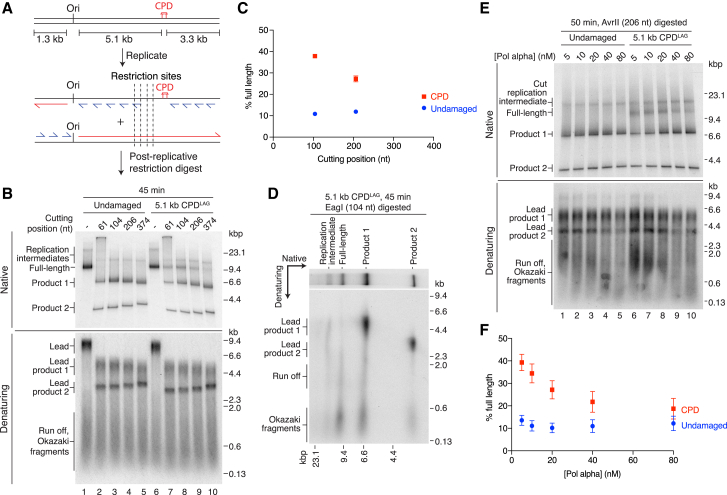
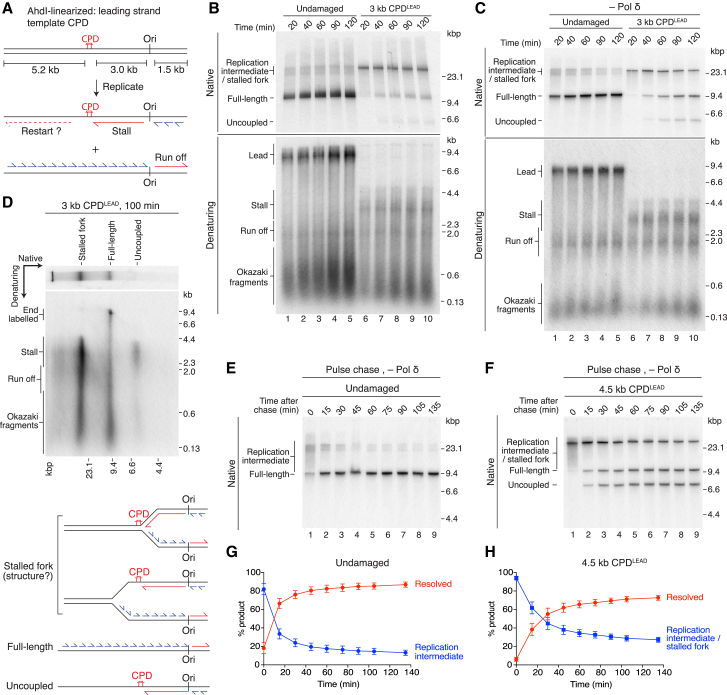
The replisome must overcome DNA damage to ensure complete chromosome replication. Here, we describe the earliest events in this process by reconstituting collisions between a eukaryotic replisome, assembled with purified proteins, and DNA damage. Lagging-strand lesions are bypassed without delay, leaving daughter-strand gaps roughly the size of an Okazaki fragment. In contrast, leading-strand polymerase stalling significantly impacts replication fork progression. We reveal that the core replisome itself can bypass leading-strand damage by re-priming synthesis beyond it. Surprisingly, this restart activity is rare, mainly due to inefficient leading-strand re-priming, rather than single-stranded DNA exposure or primer extension. We find several unanticipated mechanistic distinctions between leading- and lagging-strand priming that we propose control the replisome's initial response to DNA damage. Notably, leading-strand restart was specifically stimulated by RPA depletion, which can occur under conditions of replication stress. Our results have implications for pathway choice at stalled forks and priming at DNA replication origins.
Keywords: DNA; DNA damage response; DNA damage tolerance; DNA repair; DNA replication; genome stability; primase; re-priming; replication fork; replisome.
Copyright © 2018 MRC Laboratory of Molecular Biology. Published by Elsevier Inc. All rights reserved.
Figures
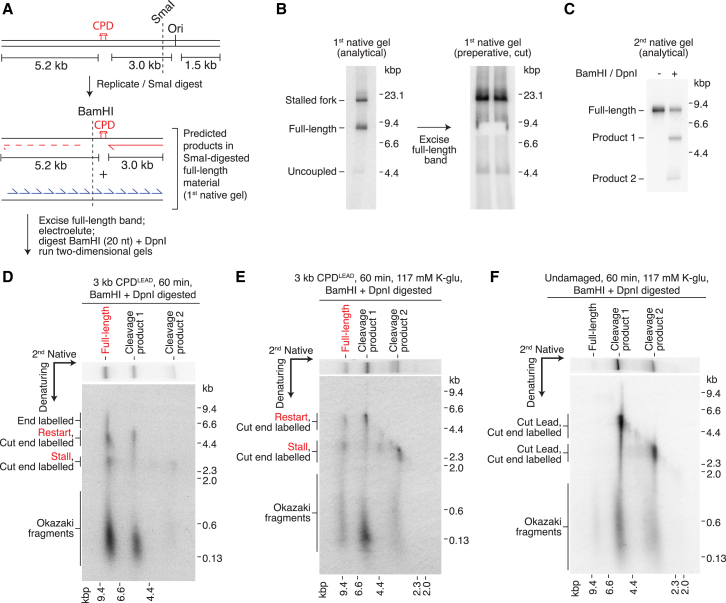
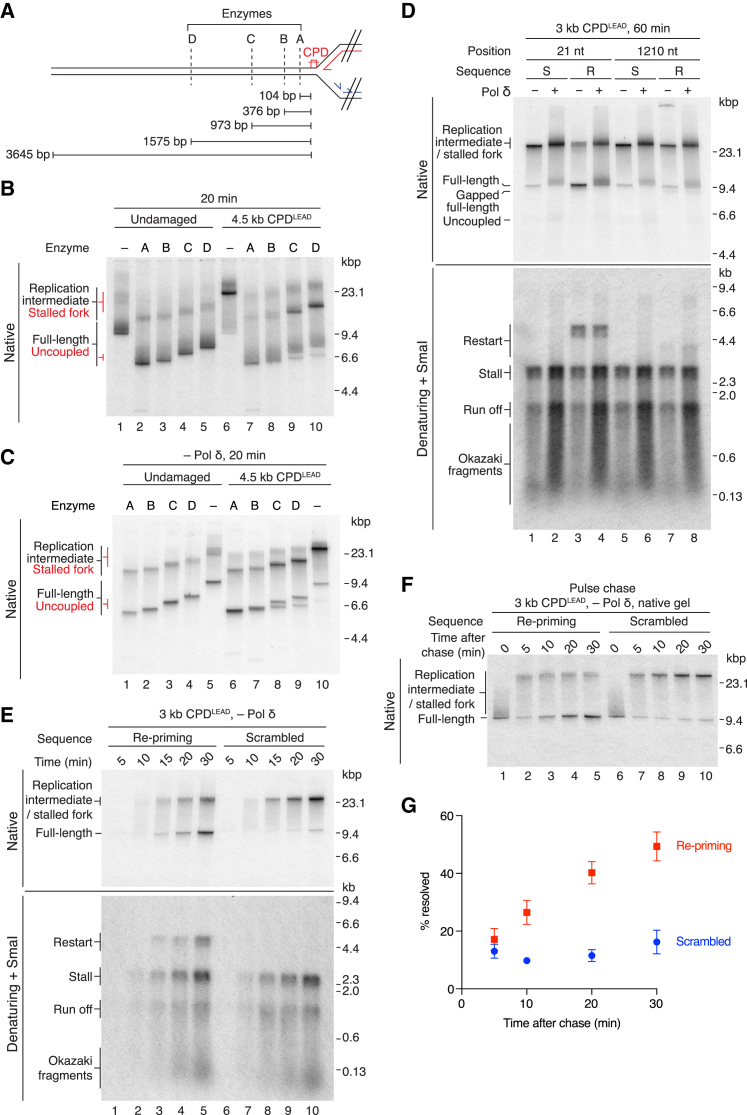
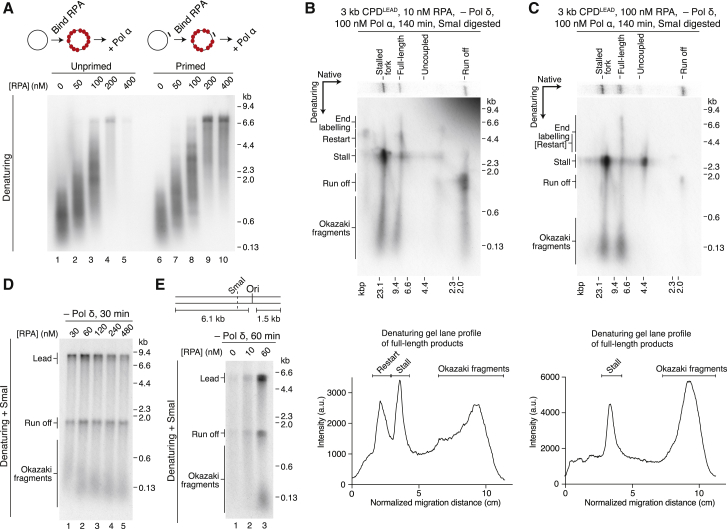
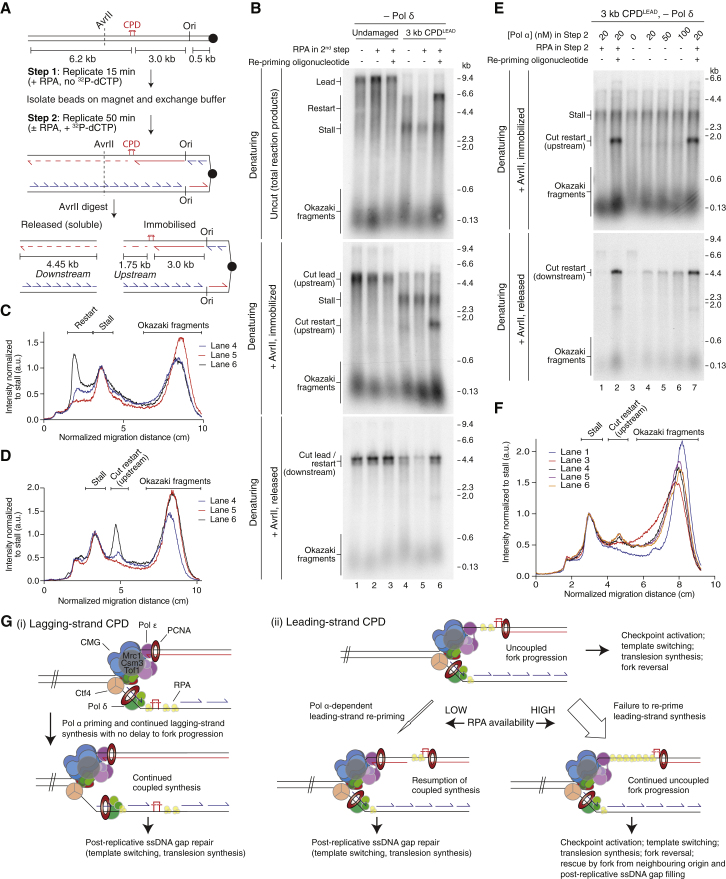
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
