

EED, a member of the polycomb group, is required for nephron differentiation and the maintenance of nephron progenitor cells
- PMID: 29945864
- PMCID: PMC6514390
- DOI: 10.1242/dev.157149
EED, a member of the polycomb group, is required for nephron differentiation and the maintenance of nephron progenitor cells
Abstract
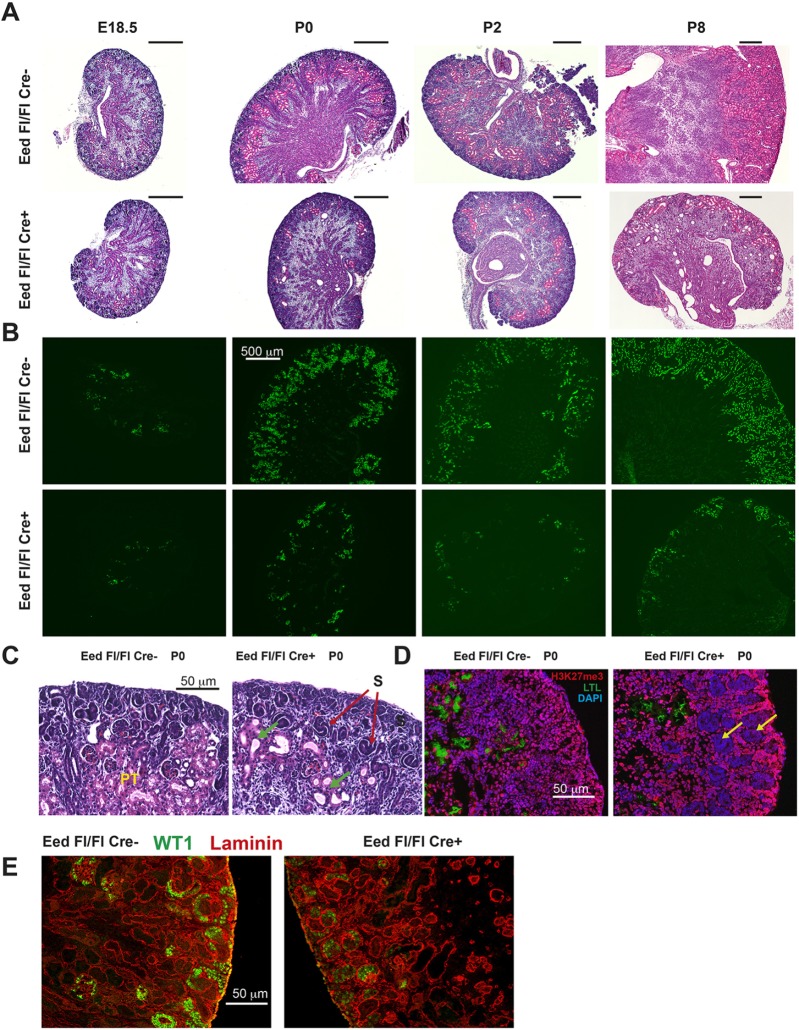
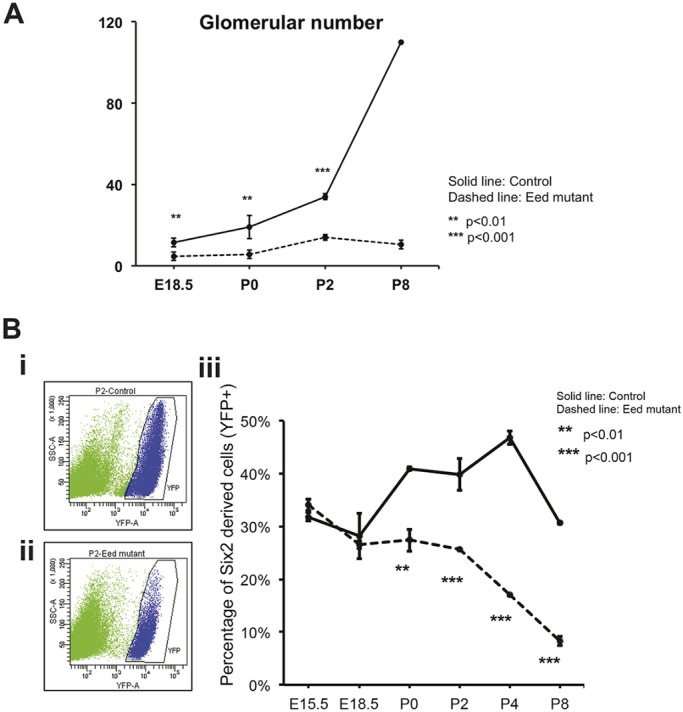
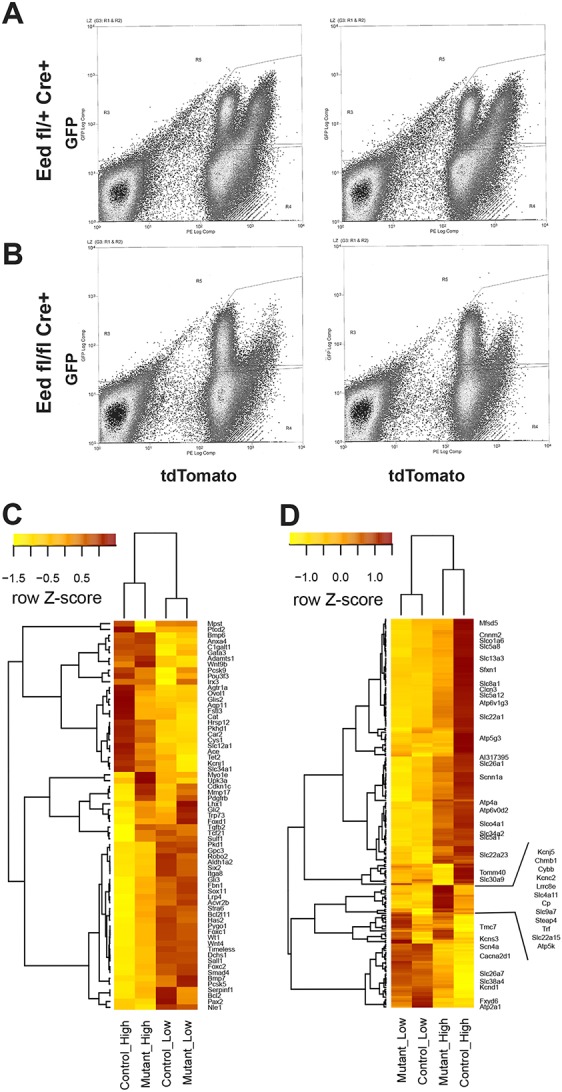
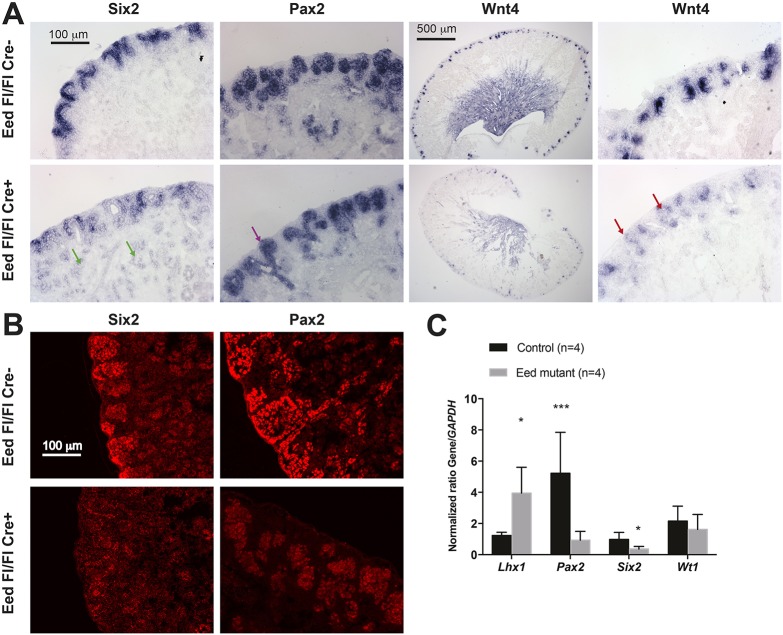
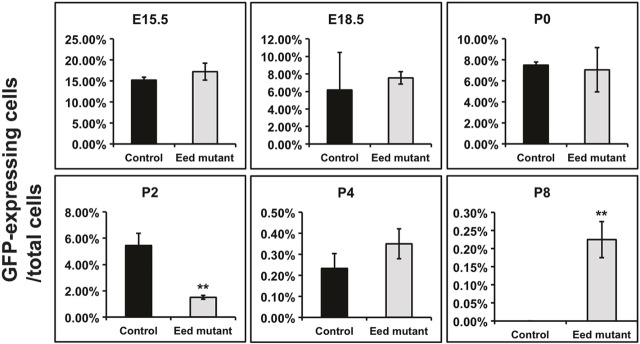
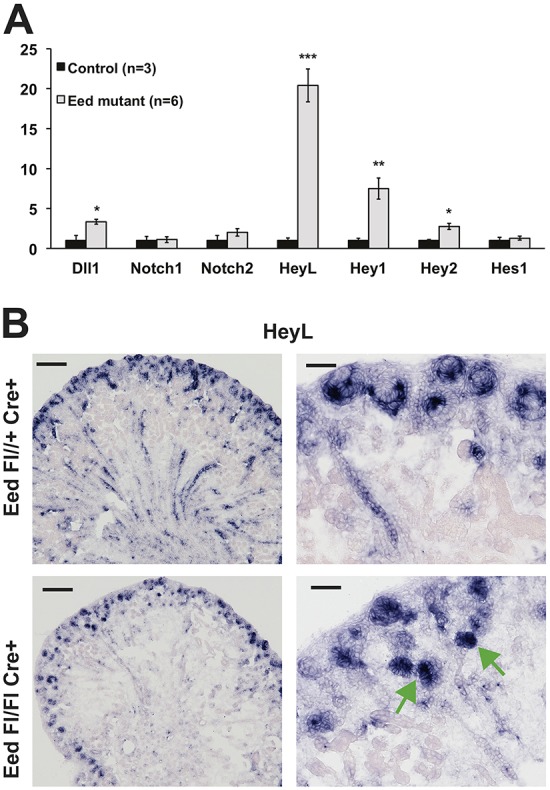
Epigenetic regulation of gene expression has a crucial role allowing for the self-renewal and differentiation of stem and progenitor populations during organogenesis. The mammalian kidney maintains a population of self-renewing stem cells that differentiate to give rise to thousands of nephrons, which are the functional units that carry out filtration to maintain physiological homeostasis. The polycomb repressive complex 2 (PRC2) epigenetically represses gene expression during development by placing the H3K27me3 mark on histone H3 at promoter and enhancer sites, resulting in gene silencing. To understand the role of PRC2 in nephron differentiation, we conditionally inactivated the Eed gene, which encodes a nonredundant component of the PRC2 complex, in nephron progenitor cells. Resultant kidneys were smaller and showed premature loss of progenitor cells. The progenitors in Eed mutant mice that were induced to differentiate did not develop into properly formed nephrons. Lhx1, normally expressed in the renal vesicle, was overexpressed in kidneys of Eed mutant mice. Thus, PRC2 has a crucial role in suppressing the expression of genes that maintain the progenitor state, allowing nephron differentiation to proceed.
Keywords: Epigenetics; Histone methylation; Kidney development; Nephron; Stem cell.
© 2018. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures
References
-
- Aiden A. P., Rivera M. N., Rheinbay E., Ku M., Coffman E. J., Truong T. T., Vargas S. O., Lander E. S., Haber D. A. and Bernstein B. E. (2010). Wilms tumor chromatin profiles highlight stem cell properties and a renal developmental network. Cell Stem Cell 6, 591-602. 10.1016/j.stem.2010.03.016 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
