

Actin filaments partition primary cilia membranes into distinct fluid corrals
- PMID: 29945903
- PMCID: PMC6080922
- DOI: 10.1083/jcb.201711104
Actin filaments partition primary cilia membranes into distinct fluid corrals
Abstract
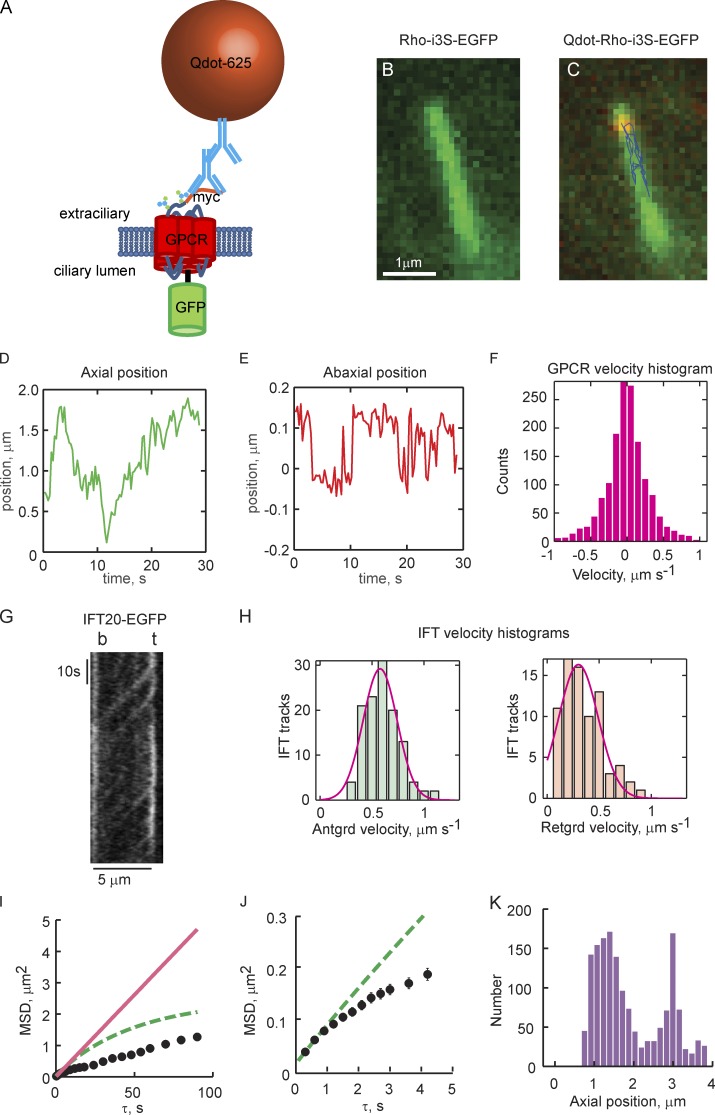
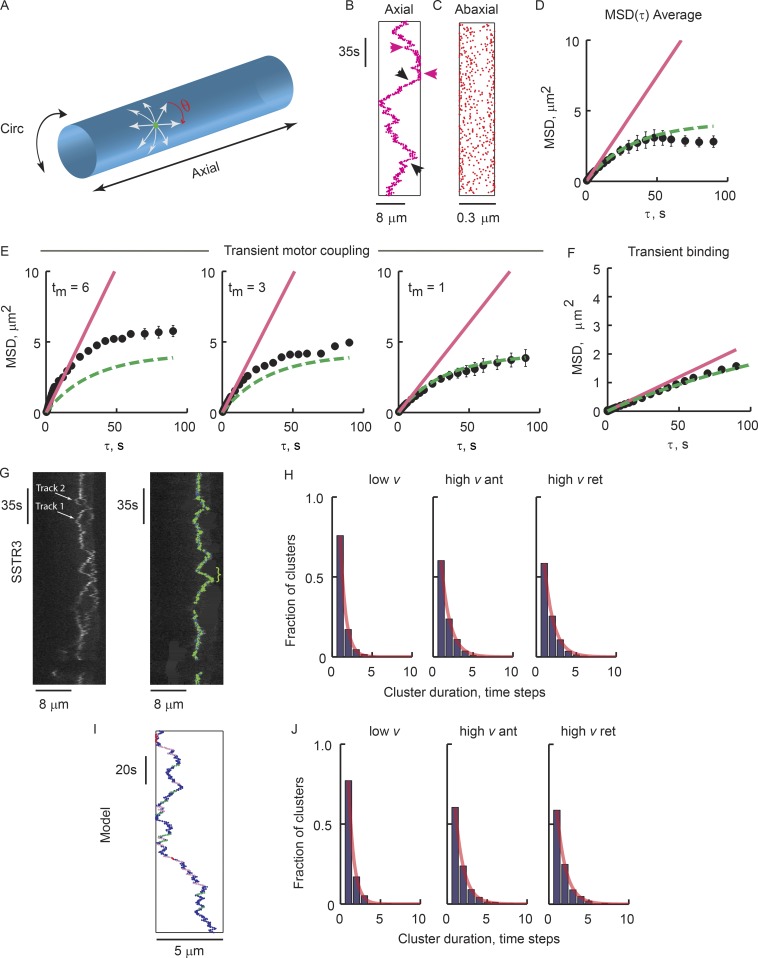
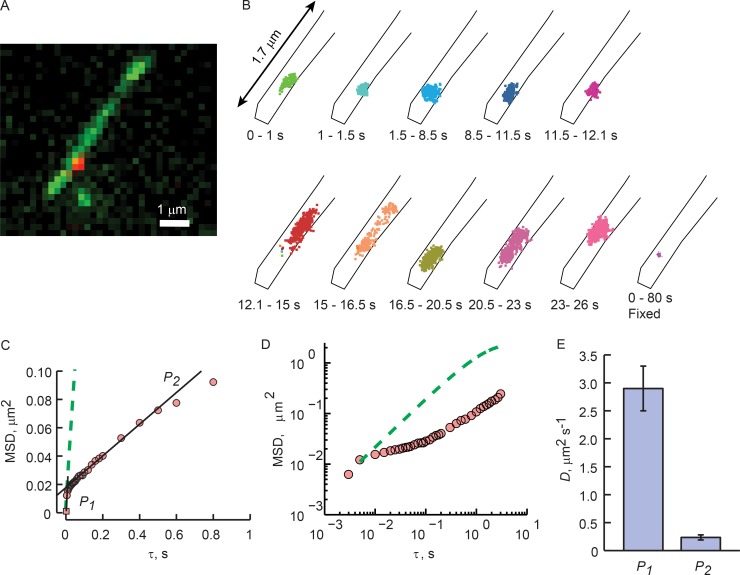
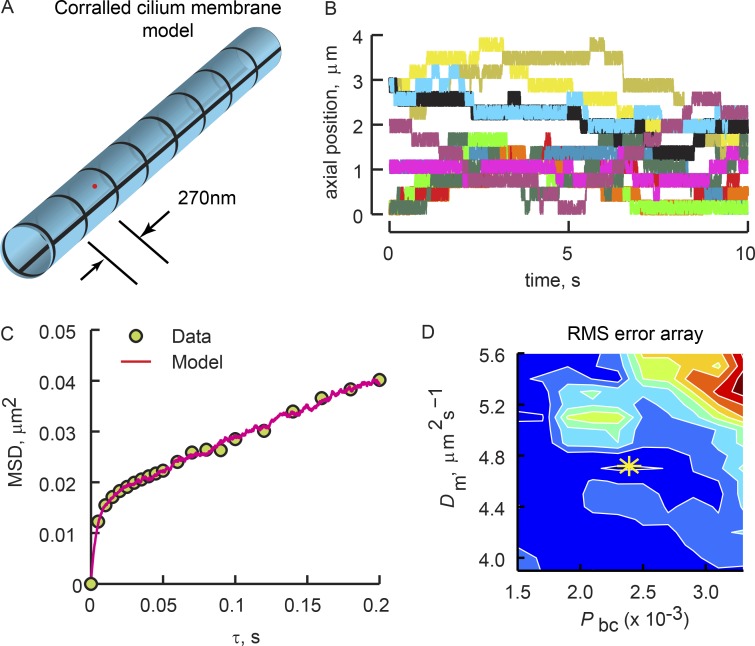
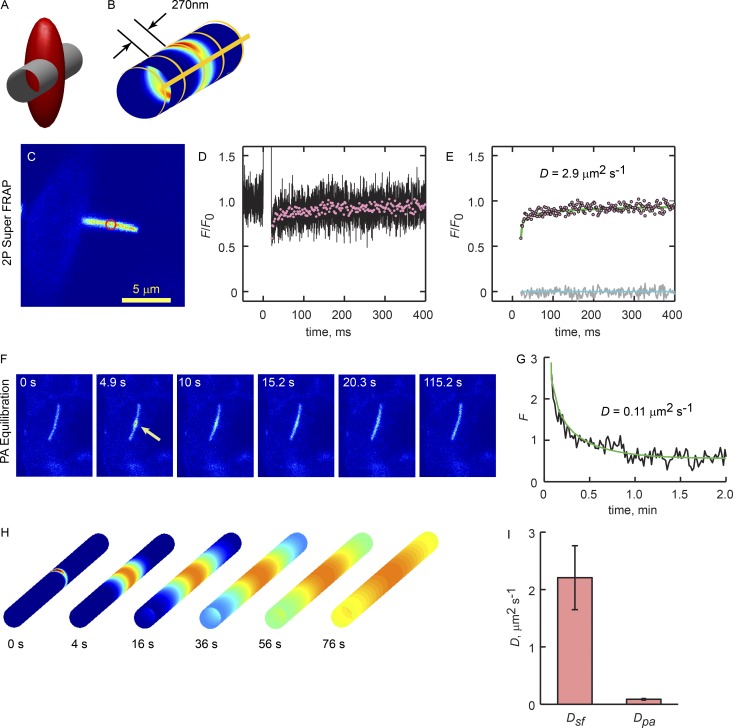
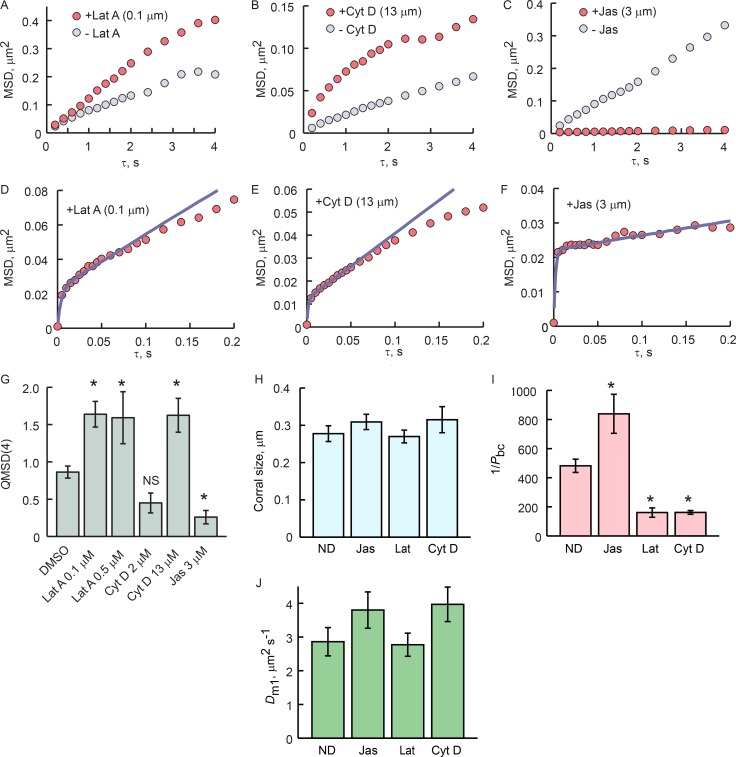
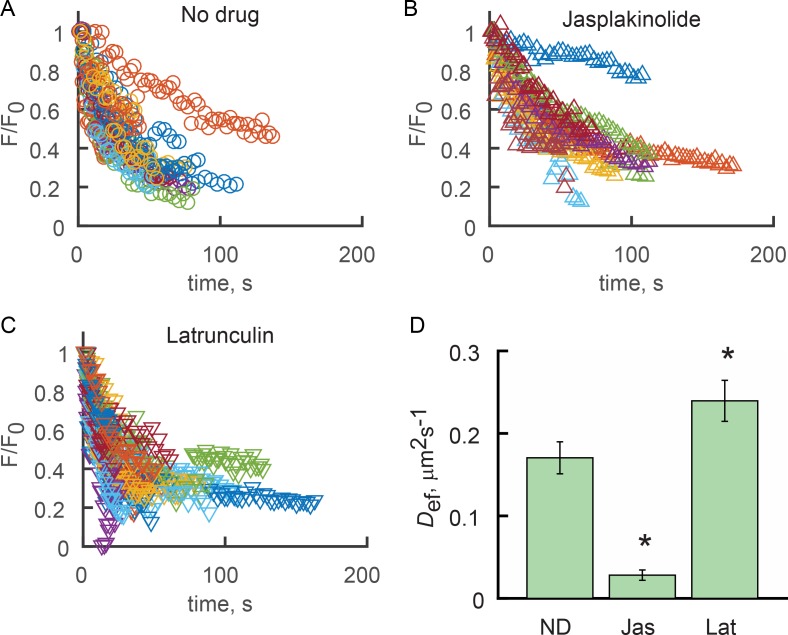
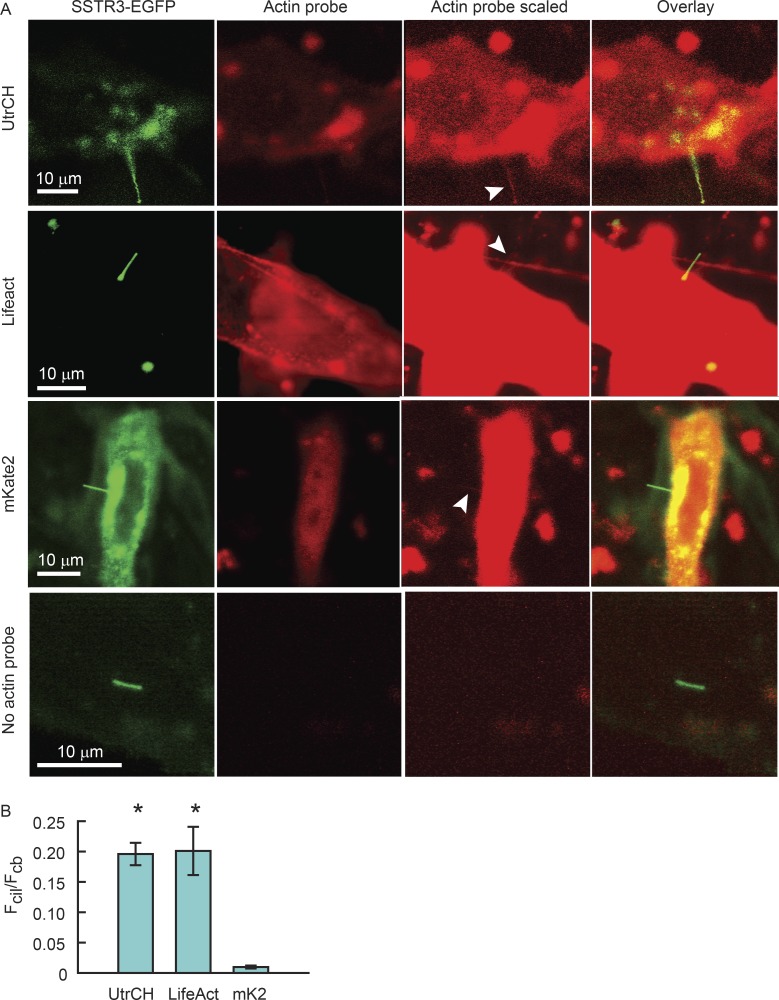
Physical properties of primary cilia membranes in living cells were examined using two independent, high-spatiotemporal-resolution approaches: fast tracking of single quantum dot-labeled G protein-coupled receptors and a novel two-photon super-resolution fluorescence recovery after photobleaching of protein ensemble. Both approaches demonstrated the cilium membrane to be partitioned into corralled domains spanning 274 ± 20 nm, within which the receptors are transiently confined for 0.71 ± 0.09 s. The mean membrane diffusion coefficient within the corrals, Dm1 = 2.9 ± 0.41 µm2/s, showed that the ciliary membranes were among the most fluid encountered. At longer times, the apparent membrane diffusion coefficient, Dm2 = 0.23 ± 0.05 µm2/s, showed that corral boundaries impeded receptor diffusion 13-fold. Mathematical simulations predict the probability of G protein-coupled receptors crossing corral boundaries to be 1 in 472. Remarkably, latrunculin A, cytochalasin D, and jasplakinolide treatments altered the corral permeability. Ciliary membranes are thus partitioned into highly fluid membrane nanodomains that are delimited by filamentous actin.
© 2018 Lee et al.
Figures
References
-
- Bouchaud J.-P., and Georges A.. 1990. Anomalous diffusion in disordered media: Statistical mechanisms, models and physical applications. Phys. Rep. 195:127–293. 10.1016/0370-1573(90)90099-N - DOI
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
