

Quantifying and Localizing the Mitochondrial Proteome Across Five Tissues in A Mouse Population
- PMID: 29945935
- PMCID: PMC6126393
- DOI: 10.1074/mcp.RA118.000554
Quantifying and Localizing the Mitochondrial Proteome Across Five Tissues in A Mouse Population
Abstract
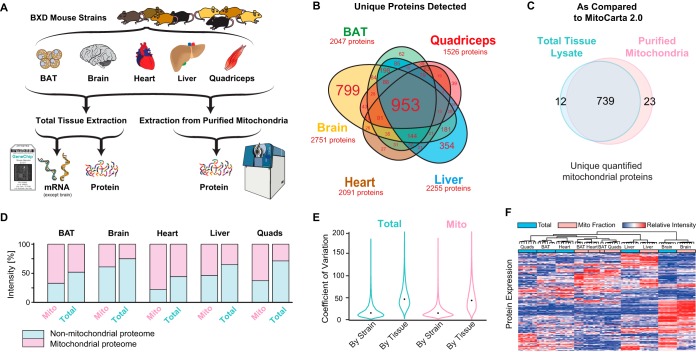
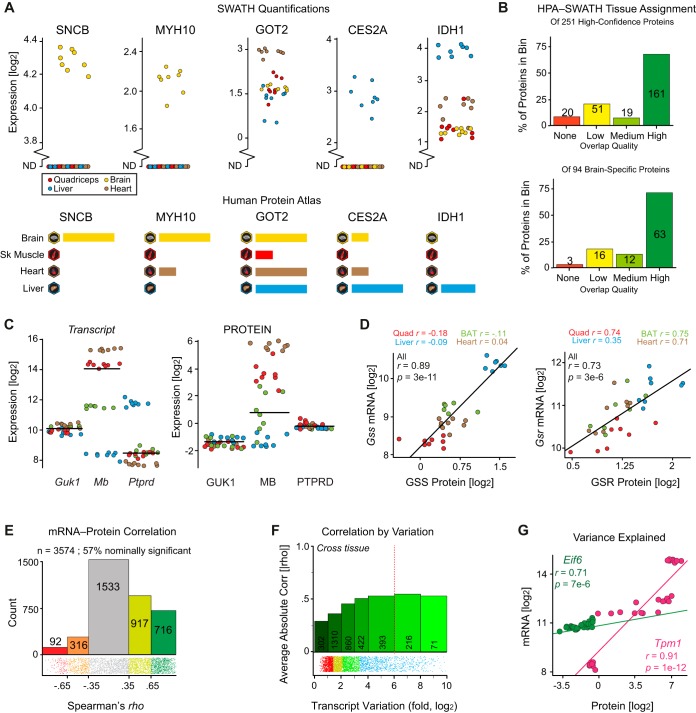
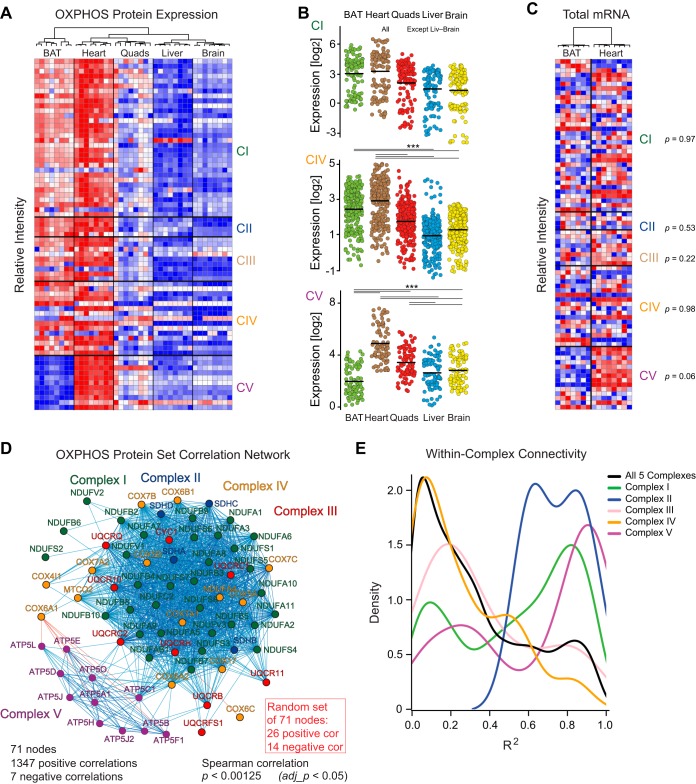
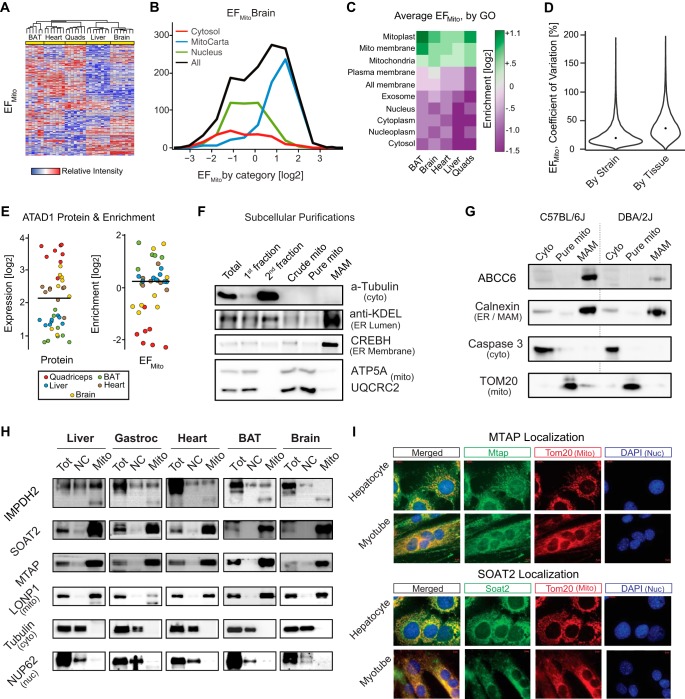
We have used SWATH mass spectrometry to quantify 3648 proteins across 76 proteomes collected from genetically diverse BXD mouse strains in two fractions (mitochondria and total cell) from five tissues: liver, quadriceps, heart, brain, and brown adipose (BAT). Across tissues, expression covariation between genes' proteins and transcripts-measured in the same individuals-broadly aligned. Covariation was however far stronger in certain subsets than others: only 8% of transcripts in the lowest expression and variance quintile covaried with their protein, in contrast to 65% of transcripts in the highest quintiles. Key functional differences among the 3648 genes were also observed across tissues, with electron transport chain (ETC) genes particularly investigated. ETC complex proteins covary and form strong gene networks according to tissue, but their equivalent transcripts do not. Certain physiological consequences, such as the depletion of ATP synthase in BAT, are thus obscured in transcript data. Lastly, we compared the quantitative proteomic measurements between the total cell and mitochondrial fractions for the five tissues. The resulting enrichment score highlighted several hundred proteins which were strongly enriched in mitochondria, which included several dozen proteins were not reported in literature to be mitochondrially localized. Four of these candidates were selected for biochemical validation, where we found MTAP, SOAT2, and IMPDH2 to be localized inside the mitochondria, whereas ABCC6 was in the mitochondria-associated membrane. These findings demonstrate the synergies of a multi-omics approach to study complex metabolic processes, and this provides a resource for further discovery and analysis of proteoforms, modified proteins, and protein localization.
© 2018 Williams et al.
Figures
References
-
- Neupert W., and Herrmann J. M. (2007) Translocation of proteins into mitochondria. Annu. Rev. Biochem. 76, 723–749 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
