

Sex differences in neural mechanisms mediating reward and addiction
- PMID: 29946108
- PMCID: PMC6235836
- DOI: 10.1038/s41386-018-0125-6
Sex differences in neural mechanisms mediating reward and addiction
Abstract
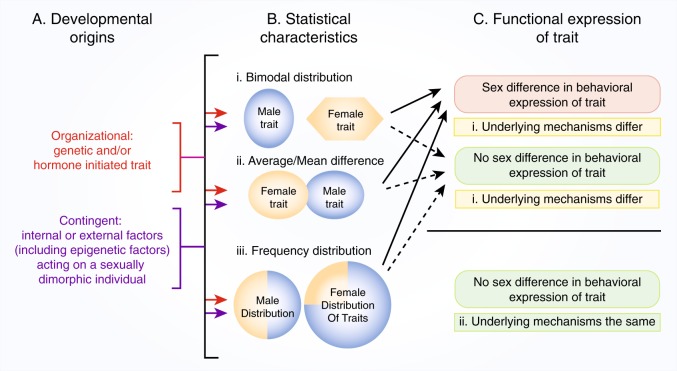
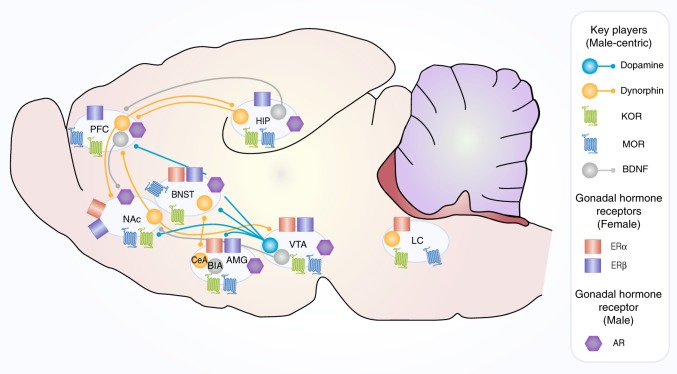
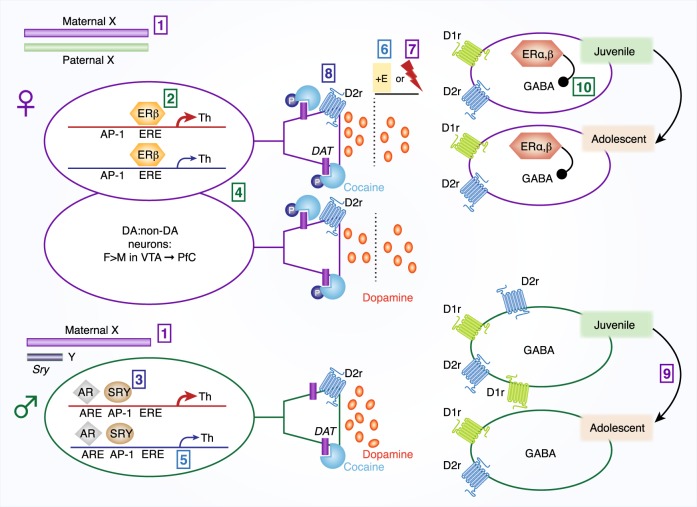
There is increasing evidence in humans and laboratory animals for biologically based sex differences in every phase of drug addiction: acute reinforcing effects, transition from occasional to compulsive use, withdrawal-associated negative affective states, craving, and relapse. There is also evidence that many qualitative aspects of the addiction phases do not differ significantly between males and females, but one sex may be more likely to exhibit a trait than the other, resulting in population differences. The conceptual framework of this review is to focus on hormonal, chromosomal, and epigenetic organizational and contingent, sex-dependent mechanisms of four neural systems that are known-primarily in males-to be key players in addiction: dopamine, mu-opioid receptors (MOR), kappa opioid receptors (KOR), and brain-derived neurotrophic factor (BDNF). We highlight data demonstrating sex differences in development, expression, and function of these neural systems as they relate-directly or indirectly-to processes of reward and addictive behavior, with a focus on psychostimulants and opioids. We identify gaps in knowledge about how these neural systems interact with sex to influence addictive behavior, emphasizing throughout that the impact of sex can be highly nuanced and male/female data should be reported regardless of the outcome.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures

References
-
- Wagner FA, Anthony JC. From first drug use to drug dependence; developmental periods of risk for dependence upon marijuana, cocaine, and alcohol. Neuropsychopharmacology. 2002;26:479–88. - PubMed
-
- Wagner FA, Anthony JC. Male-female differences in the risk of progression from first use to dependence upon cannabis, cocaine, and alcohol. Drug Alcohol Depend. 2007;86:191–8. - PubMed
-
- NSDUH NSoDUaH. Substance Abuse and Mental Health Services Administration, Results from the 2013 National Survey on Drug Use and Health: Summary of National Findings, NSDUH Series H-48, HHS Publication No. (SMA) 14-4863. Rockville, MD: Substance Abuse and Mental Health Services Administration, 2014.
-
- Cotto JH, Davis E, Dowling GJ, Elcano JC, Staton AB, Weiss SR. Gender effects on drug use, abuse, and dependence: a special analysis of results from the National Survey on Drug Use and Health. Gend Med. 2010;7:402–13. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
