

Structural basis for terminal loop recognition and stimulation of pri-miRNA-18a processing by hnRNP A1
- PMID: 29946118
- PMCID: PMC6018666
- DOI: 10.1038/s41467-018-04871-9
Structural basis for terminal loop recognition and stimulation of pri-miRNA-18a processing by hnRNP A1
Abstract
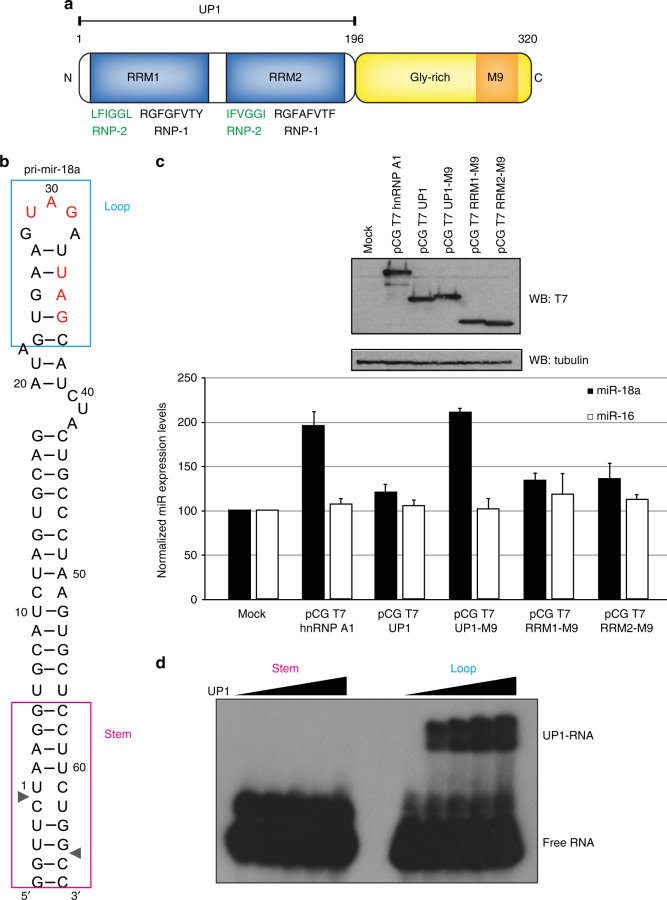
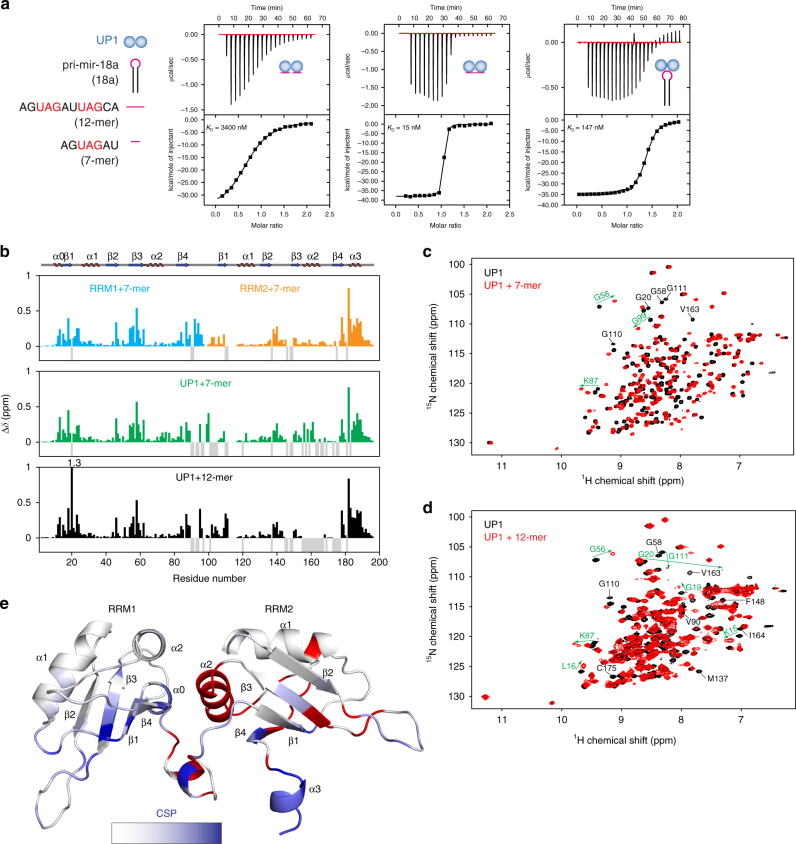
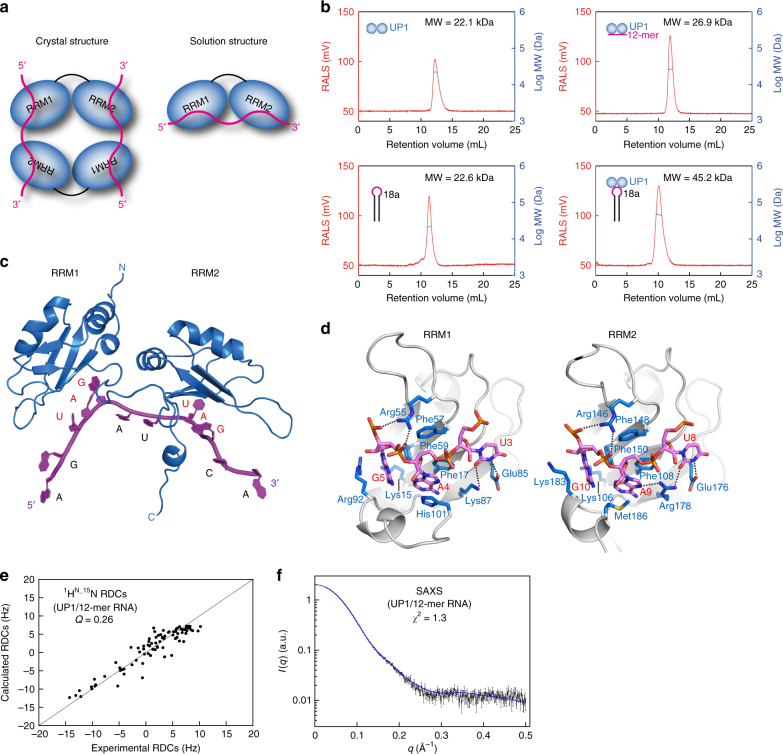
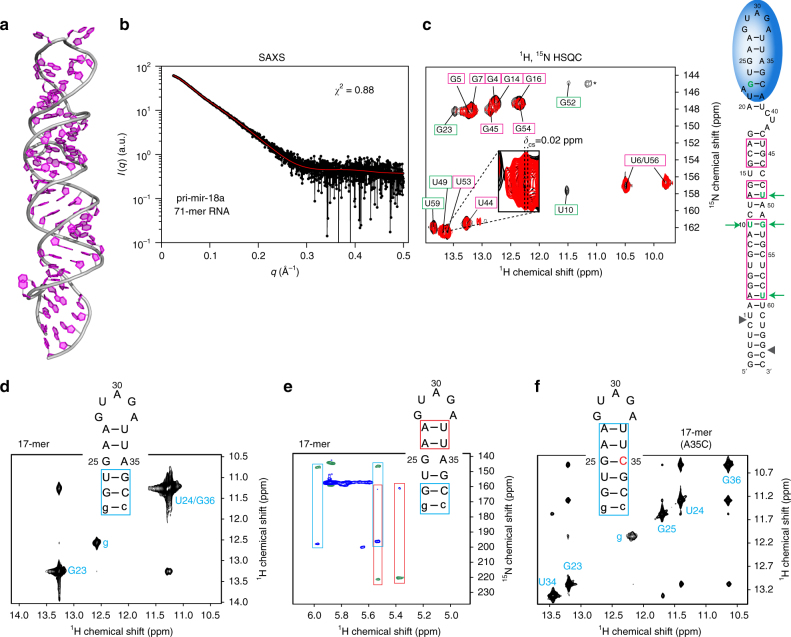
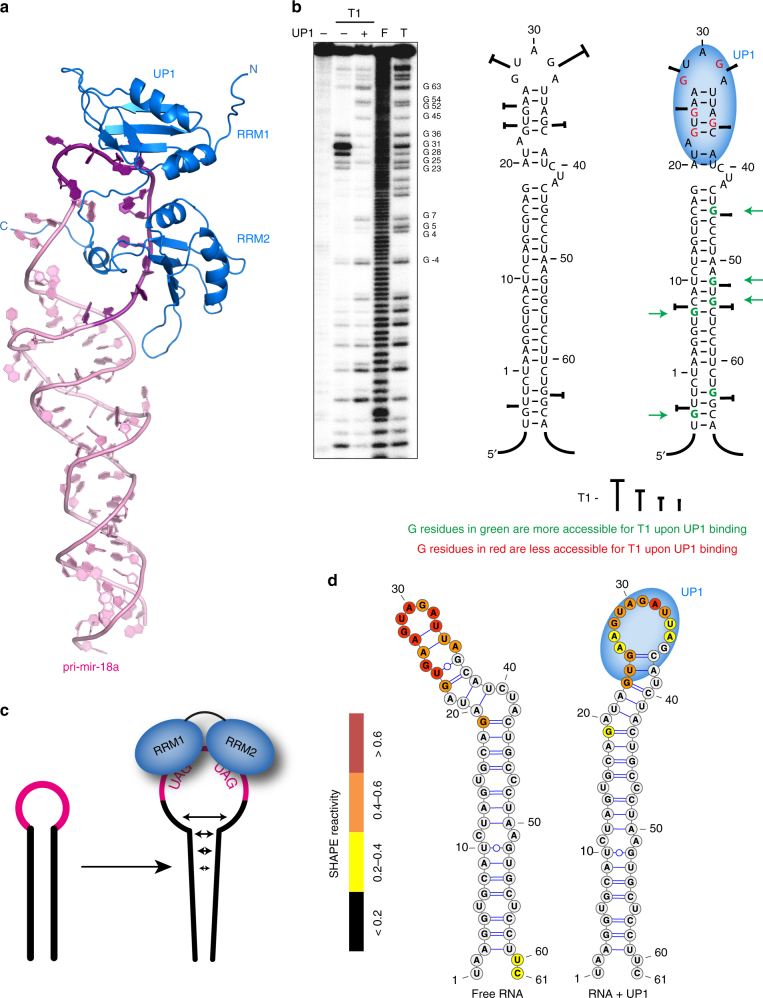
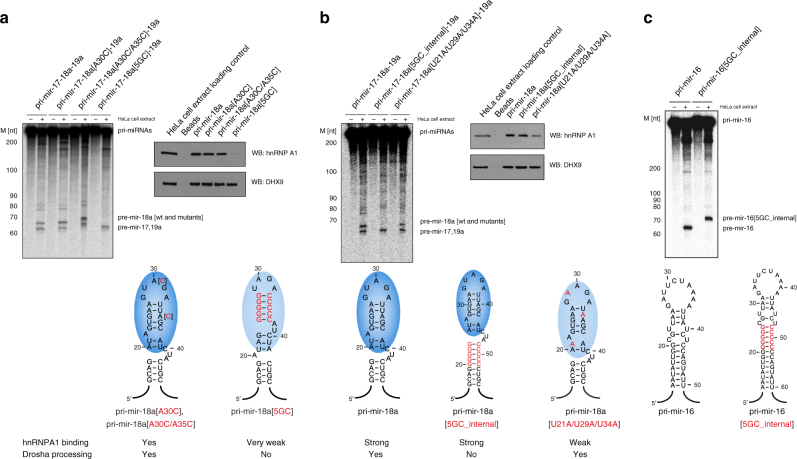
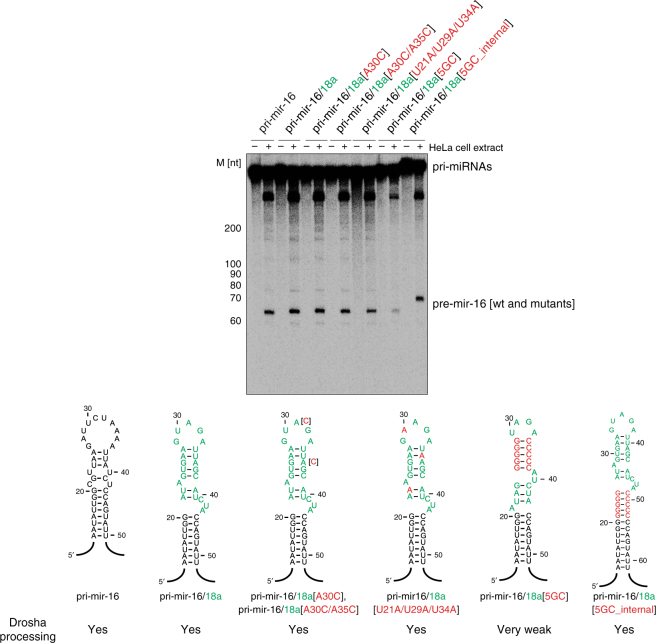
Post-transcriptional mechanisms play a predominant role in the control of microRNA (miRNA) production. Recognition of the terminal loop of precursor miRNAs by RNA-binding proteins (RBPs) influences their processing; however, the mechanistic basis for how levels of individual or subsets of miRNAs are regulated is mostly unexplored. We previously showed that hnRNP A1, an RBP implicated in many aspects of RNA processing, acts as an auxiliary factor that promotes the Microprocessor-mediated processing of pri-mir-18a. Here, by using an integrative structural biology approach, we show that hnRNP A1 forms a 1:1 complex with pri-mir-18a where both RNA recognition motifs (RRMs) bind to cognate RNA sequence motifs in the terminal loop of pri-mir-18a. Terminal loop binding induces an allosteric destabilization of base-pairing in the pri-mir-18a stem that promotes its downstream processing. Our results highlight terminal loop RNA recognition by RBPs as a potential general principle of miRNA biogenesis and regulation.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Conformational Effects of a Cancer-Linked Mutation in Pri-miR-30c RNA.J Mol Biol. 2022 Sep 30;434(18):167705. doi: 10.1016/j.jmb.2022.167705. Epub 2022 Jun 24. J Mol Biol. 2022. PMID: 35760371
-
Posttranscriptional regulation of miRNAs harboring conserved terminal loops.Mol Cell. 2008 Nov 7;32(3):383-93. doi: 10.1016/j.molcel.2008.10.013. Mol Cell. 2008. PMID: 18995836 Free PMC article.
-
The multifunctional RNA-binding protein hnRNP A1 is required for processing of miR-18a.Nat Struct Mol Biol. 2007 Jul;14(7):591-6. doi: 10.1038/nsmb1250. Epub 2007 Jun 10. Nat Struct Mol Biol. 2007. PMID: 17558416
-
Stimulation of pri-miR-18a processing by hnRNP A1.Adv Exp Med Biol. 2010;700:28-35. doi: 10.1007/978-1-4419-7823-3_3. Adv Exp Med Biol. 2010. PMID: 21627027 Review.
-
Idiosyncrasies of hnRNP A1-RNA recognition: Can binding mode influence function.Semin Cell Dev Biol. 2019 Feb;86:150-161. doi: 10.1016/j.semcdb.2018.04.001. Epub 2018 Apr 9. Semin Cell Dev Biol. 2019. PMID: 29625167 Free PMC article. Review.
Cited by
-
hnRNP A/B Proteins: An Encyclopedic Assessment of Their Roles in Homeostasis and Disease.Biology (Basel). 2021 Jul 24;10(8):712. doi: 10.3390/biology10080712. Biology (Basel). 2021. PMID: 34439945 Free PMC article. Review.
-
Unraveling the advances of non-coding RNAs on the tumor microenvironment: innovative strategies for cancer therapies.J Transl Med. 2025 Jun 2;23(1):614. doi: 10.1186/s12967-025-06629-6. J Transl Med. 2025. PMID: 40457447 Free PMC article. Review.
-
HRPK-1, a conserved KH-domain protein, modulates microRNA activity during Caenorhabditis elegans development.PLoS Genet. 2019 Oct 4;15(10):e1008067. doi: 10.1371/journal.pgen.1008067. eCollection 2019 Oct. PLoS Genet. 2019. PMID: 31584932 Free PMC article.
-
Chronic myelogenous leukemia cells remodel the bone marrow niche via exosome-mediated transfer of miR-320.Theranostics. 2019 Jul 28;9(19):5642-5656. doi: 10.7150/thno.34813. eCollection 2019. Theranostics. 2019. PMID: 31534508 Free PMC article.
-
Exploring the roles and molecular mechanisms of RNA binding proteins in the sorting of noncoding RNAs into exosomes during tumor progression.J Adv Res. 2024 Nov;65:105-123. doi: 10.1016/j.jare.2023.11.029. Epub 2023 Nov 27. J Adv Res. 2024. PMID: 38030125 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
