

Bacillus subtilis RarA modulates replication restart
- PMID: 29947798
- PMCID: PMC6101539
- DOI: 10.1093/nar/gky541
Bacillus subtilis RarA modulates replication restart
Abstract
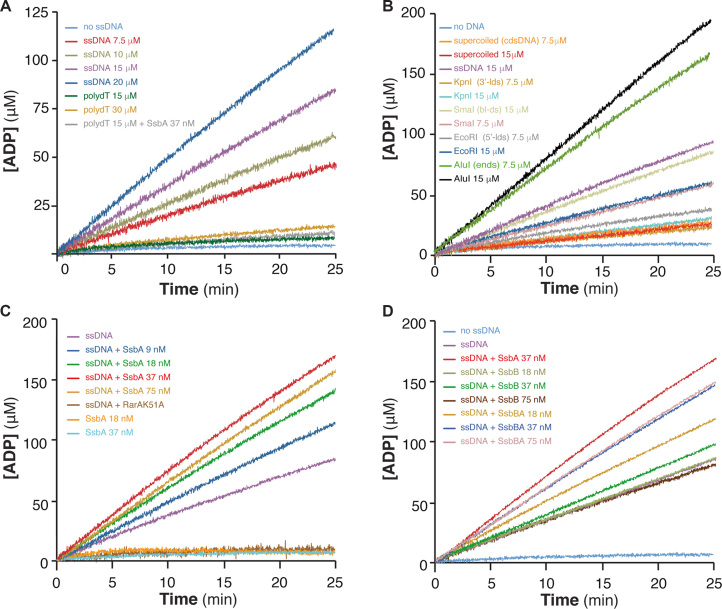
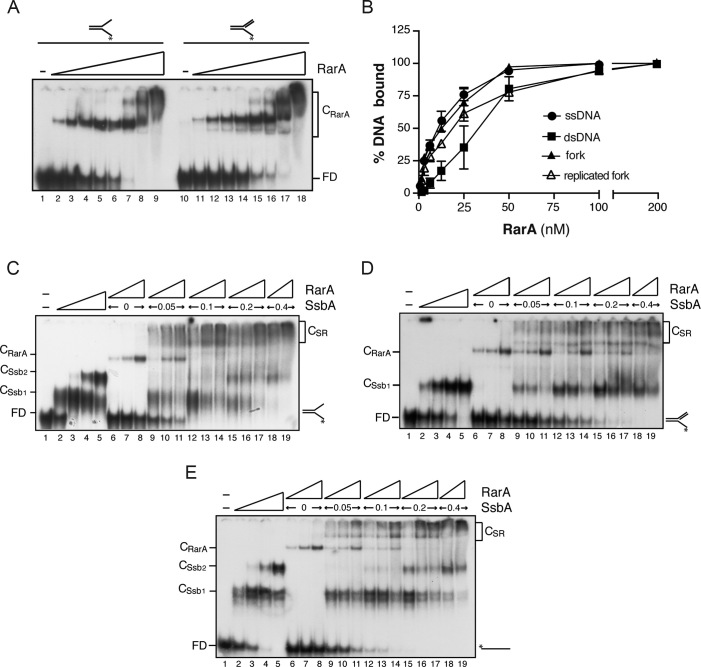
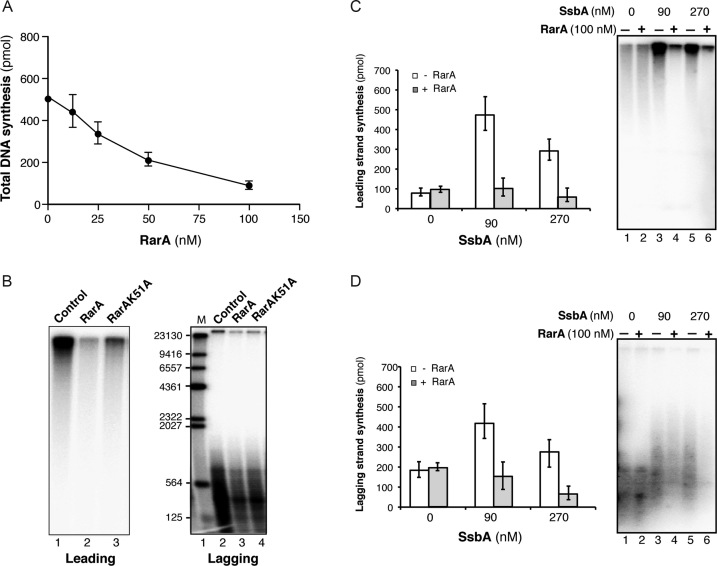
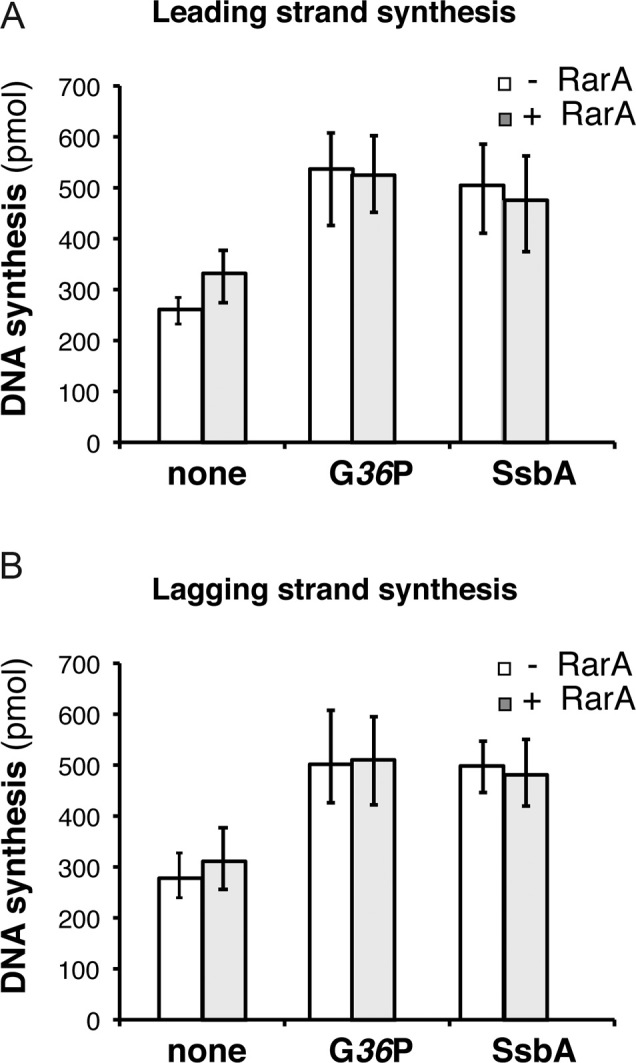
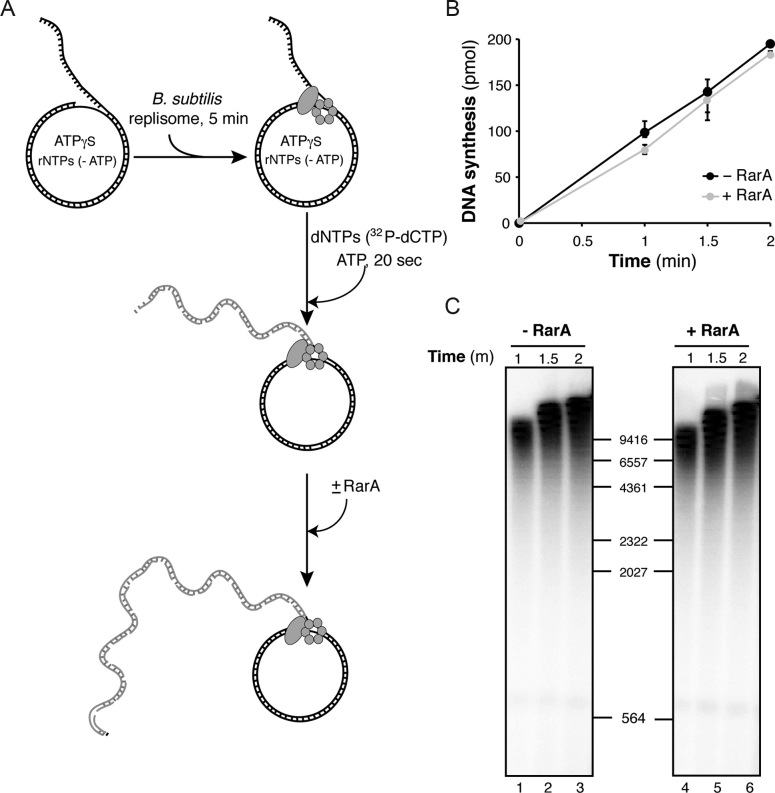
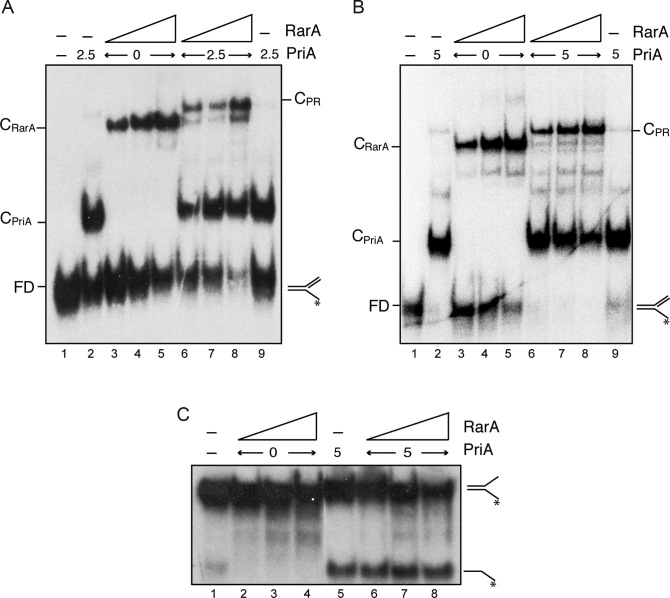
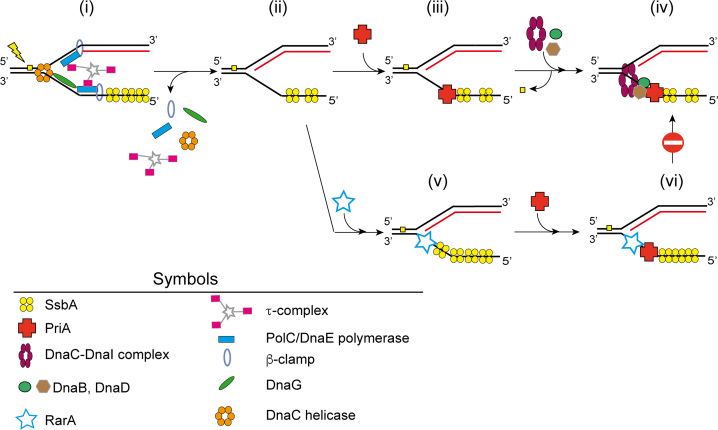
The ubiquitous RarA/Mgs1/WRNIP protein plays a crucial, but poorly understood role in genome maintenance. We show that Bacillus subtilis RarA, in the apo form, preferentially binds single-stranded (ss) over double-stranded (ds) DNA. SsbA bound to ssDNA loads RarA, and for such recruitment the amphipathic C-terminal domain of SsbA is required. RarA is a DNA-dependent ATPase strongly stimulated by ssDNA-dsDNA junctions and SsbA, or by dsDNA ends. RarA, which may interact with PriA, does not stimulate PriA DNA unwinding. In a reconstituted PriA-dependent DNA replication system, RarA inhibited initiation, but not chain elongation. The RarA effect was not observed in the absence of SsbA, or when the host-encoded preprimosome and the DNA helicase are replaced by proteins from the SPP1 phage with similar function. We propose that RarA assembles at blocked forks to maintain genome integrity. Through its interaction with SsbA and with a preprimosomal component, RarA might impede the assembly of the replicative helicase, to prevent that recombination intermediates contribute to pathological DNA replication restart.
Figures
Similar articles
-
Function of a strand-separation pin element in the PriA DNA replication restart helicase.J Biol Chem. 2019 Feb 22;294(8):2801-2814. doi: 10.1074/jbc.RA118.006870. Epub 2018 Dec 28. J Biol Chem. 2019. PMID: 30593500 Free PMC article.
-
Restart of DNA replication in Gram-positive bacteria: functional characterisation of the Bacillus subtilis PriA initiator.Nucleic Acids Res. 2002 Apr 1;30(7):1593-605. doi: 10.1093/nar/30.7.1593. Nucleic Acids Res. 2002. PMID: 11917020 Free PMC article.
-
A bacterial PriB with weak single-stranded DNA binding activity can stimulate the DNA unwinding activity of its cognate PriA helicase.BMC Microbiol. 2011 Aug 23;11:189. doi: 10.1186/1471-2180-11-189. BMC Microbiol. 2011. PMID: 21861872 Free PMC article.
-
Mechanisms of bacterial DNA replication restart.Nucleic Acids Res. 2018 Jan 25;46(2):504-519. doi: 10.1093/nar/gkx1203. Nucleic Acids Res. 2018. PMID: 29202195 Free PMC article. Review.
-
Processing of stalled replication forks in Bacillus subtilis.FEMS Microbiol Rev. 2024 Jan 12;48(1):fuad065. doi: 10.1093/femsre/fuad065. FEMS Microbiol Rev. 2024. PMID: 38052445 Free PMC article. Review.
Cited by
-
Bacillus subtilis RarA forms damage-inducible foci that scan the entire cell.BMC Res Notes. 2019 Apr 11;12(1):219. doi: 10.1186/s13104-019-4252-x. BMC Res Notes. 2019. PMID: 30971308 Free PMC article.
-
Single molecule tracking reveals functions for RarA at replication forks but also independently from replication during DNA repair in Bacillus subtilis.Sci Rep. 2019 Feb 13;9(1):1997. doi: 10.1038/s41598-018-38289-6. Sci Rep. 2019. PMID: 30760776 Free PMC article.
-
Insights into 6S RNA in lactic acid bacteria (LAB).BMC Genom Data. 2021 Sep 3;22(1):29. doi: 10.1186/s12863-021-00983-2. BMC Genom Data. 2021. PMID: 34479493 Free PMC article.
-
Bacillus subtilis PcrA Couples DNA Replication, Transcription, Recombination and Segregation.Front Mol Biosci. 2020 Jul 21;7:140. doi: 10.3389/fmolb.2020.00140. eCollection 2020. Front Mol Biosci. 2020. PMID: 32793628 Free PMC article.
-
Crystal Structure of DNA Replication Protein SsbA Complexed with the Anticancer Drug 5-Fluorouracil.Int J Mol Sci. 2023 Oct 4;24(19):14899. doi: 10.3390/ijms241914899. Int J Mol Sci. 2023. PMID: 37834349 Free PMC article.
References
-
- Lau I.F., Filipe S.R., Soballe B., Okstad O.A., Barre F.X., Sherratt D.J.. Spatial and temporal organization of replicating Escherichia coli chromosomes. Mol. Microbiol. 2003; 49:731–743. - PubMed
-
- Shibata T., Hishida T., Kubota Y., Han Y.W., Iwasaki H., Shinagawa H.. Functional overlap between RecA and MgsA (RarA) in the rescue of stalled replication forks in Escherichia coli. Genes Cells. 2005; 10:181–191. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
