

The cellular machineries responsible for the division of endosymbiotic organelles
- PMID: 29948488
- PMCID: PMC6424925
- DOI: 10.1007/s10265-018-1050-9
The cellular machineries responsible for the division of endosymbiotic organelles
Erratum in
-
Correction to: The cellular machineries responsible for the division of endosymbiotic organelles.J Plant Res. 2019 Mar;132(2):297-298. doi: 10.1007/s10265-019-01093-y. J Plant Res. 2019. PMID: 30805737 Free PMC article.
Abstract
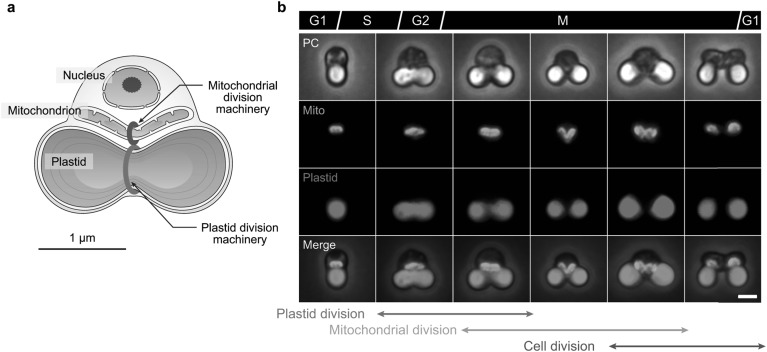
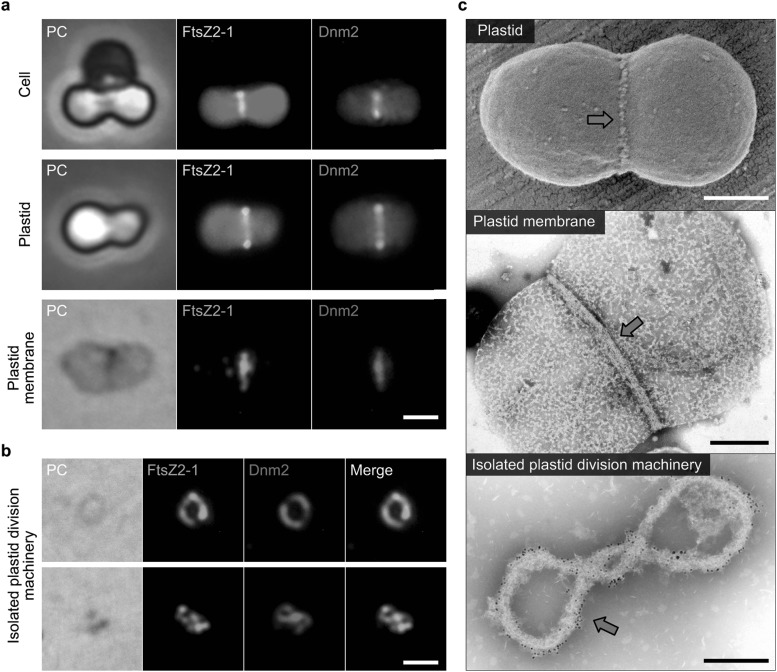
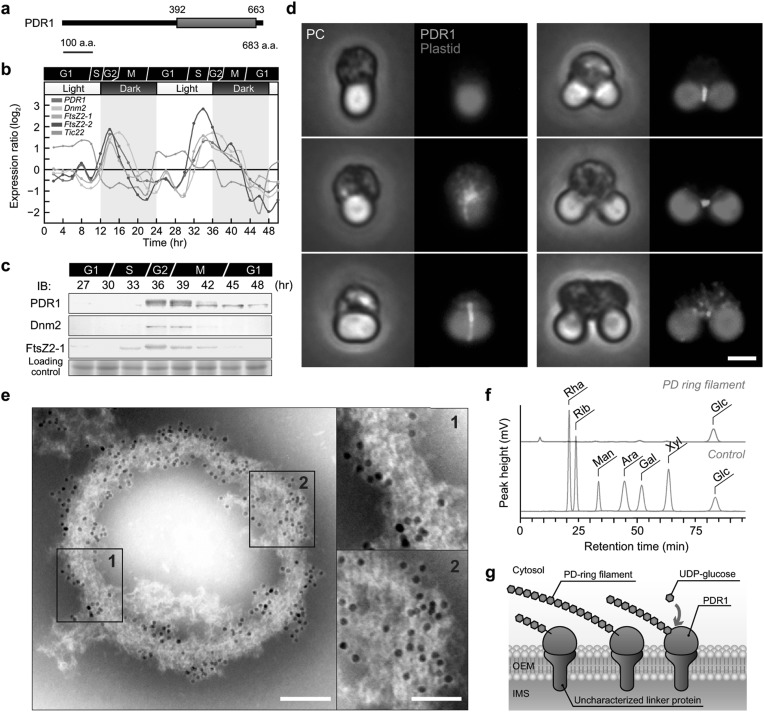
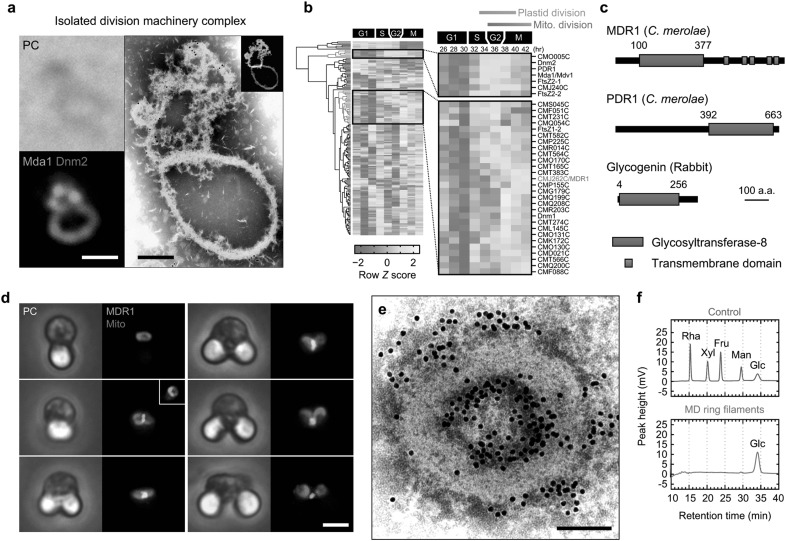
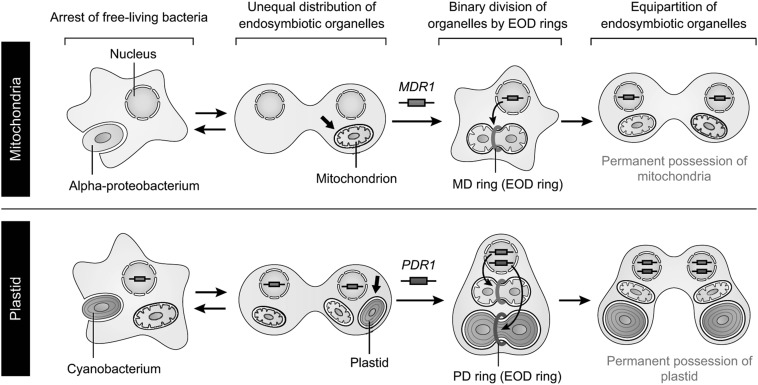
Chloroplasts (plastids) and mitochondria evolved from endosymbiotic bacteria. These organelles perform vital functions in photosynthetic eukaryotes, such as harvesting and converting energy for use in biological processes. Consistent with their evolutionary origins, plastids and mitochondria proliferate by the binary fission of pre-existing organelles. Here, I review the structures and functions of the supramolecular machineries driving plastid and mitochondrial division, which were discovered and first studied in the primitive red alga Cyanidioschyzon merolae. In the past decade, intact division machineries have been isolated from plastids and mitochondria and examined to investigate their underlying structure and molecular mechanisms. A series of studies has elucidated how these division machineries assemble and transform during the fission of these organelles, and which of the component proteins generate the motive force for their contraction. Plastid- and mitochondrial-division machineries have important similarities in their structures and mechanisms despite sharing no component proteins, implying that these division machineries evolved in parallel. The establishment of these division machineries might have enabled the host eukaryotic ancestor to permanently retain these endosymbiotic organelles by regulating their binary fission and the equal distribution of resources to daughter cells. These findings provide key insights into the establishment of endosymbiotic organelles and have opened new avenues of research into their evolution and mechanisms of proliferation.
Keywords: Chloroplast division; Endosymbiotic organelle; MDR1; Mitochondrial division; PDR1.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
